

Sulforaphane induced NRF2 activation in obese pregnancy attenuates developmental redox imbalance and improves early-life cardiovascular function in offspring
- PMID: 37774548
- PMCID: PMC10534264
- DOI: 10.1016/j.redox.2023.102883
Sulforaphane induced NRF2 activation in obese pregnancy attenuates developmental redox imbalance and improves early-life cardiovascular function in offspring
Abstract
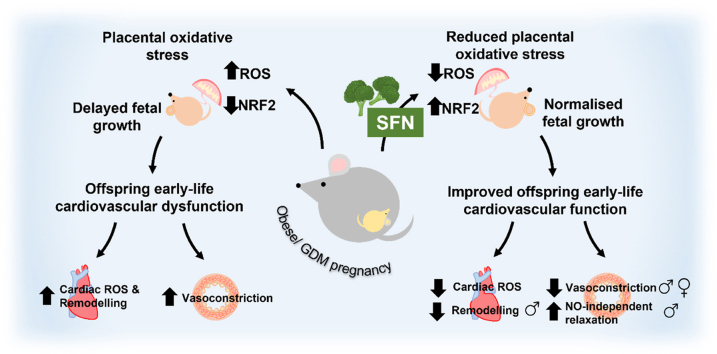
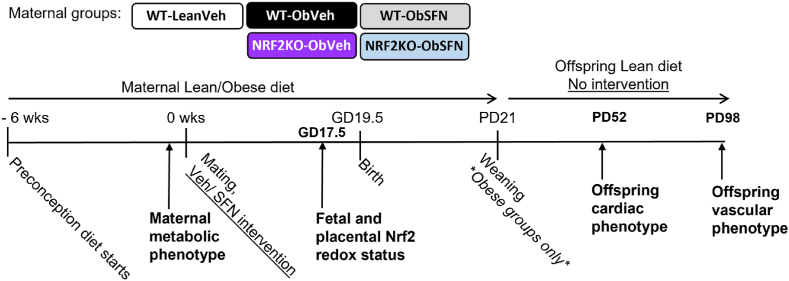
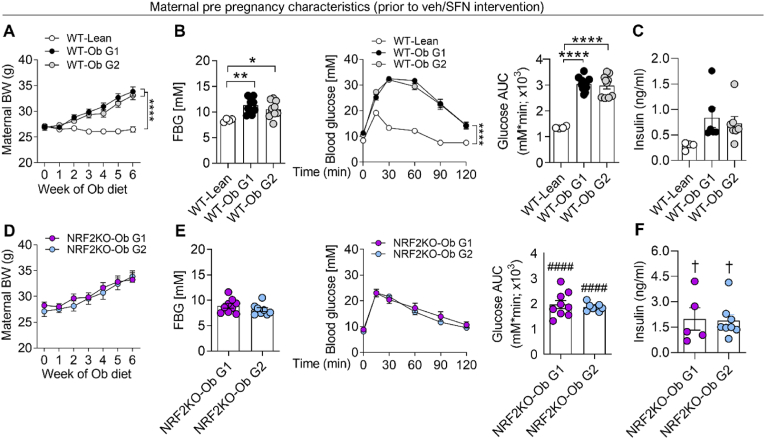
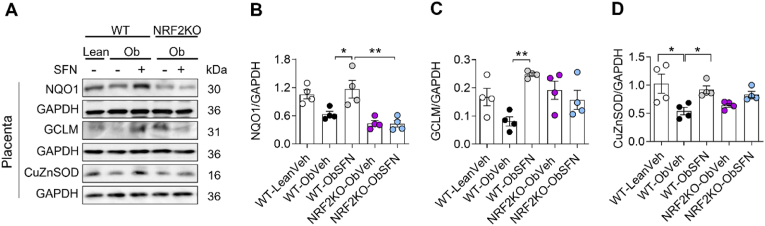
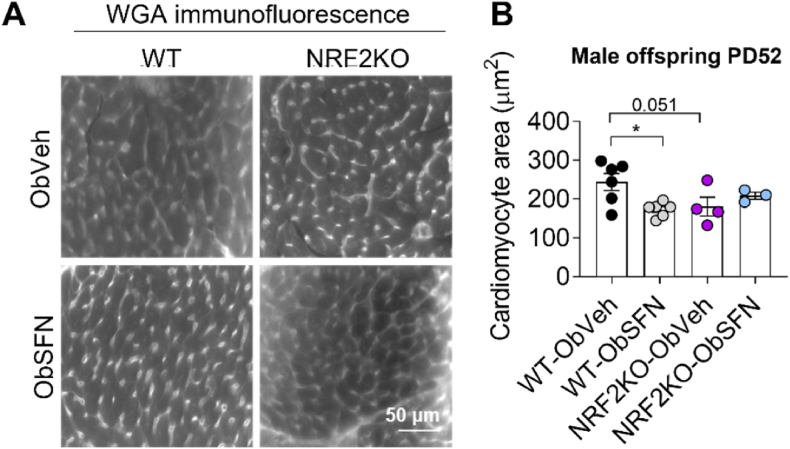
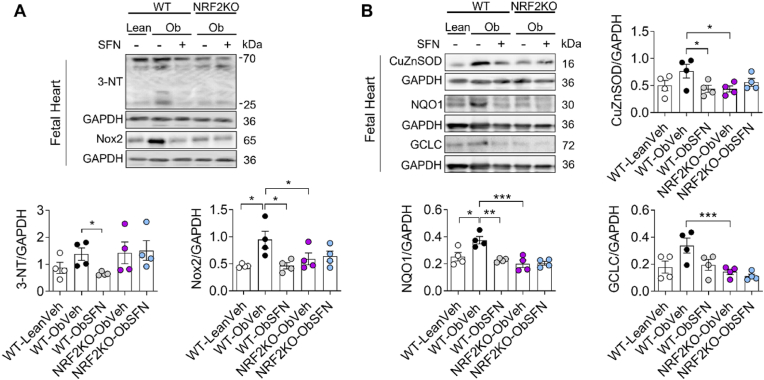
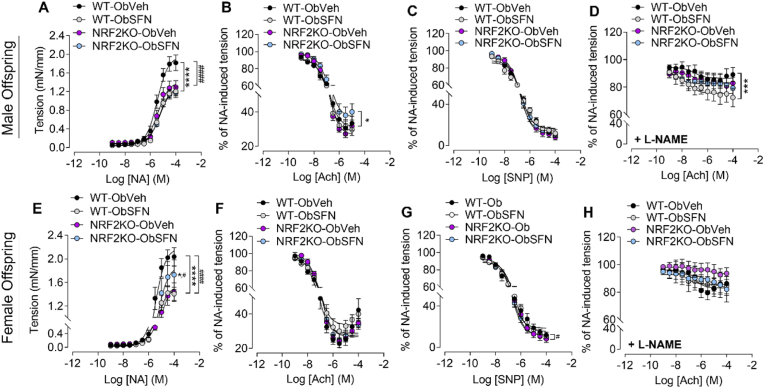
In adverse pregnancy a perturbed redox environment is associated with abnormal early-life cardiovascular development and function. Previous studies have noted alterations in the expression and/or activity of Nuclear Factor E2 Related Factor 2 (NRF2) and its antioxidant targets during human gestational diabetic (GDM) pregnancy, however to our knowledge the functional role of NRF2 in fetal 'priming' of cardiovascular dysfunction in obese and GDM pregnancy has not been investigated. Using a murine model of obesity-induced glucose dysregulated pregnancy, we demonstrate that NRF2 activation by maternal sulforaphane (SFN) supplementation normalizes NRF2-linked NQO1, GCL and CuZnSOD expression in maternal and fetal liver placental and fetal heart tissue by gestational day 17.5. Activation of NRF2 in utero in wild type but not NRF2 deficient mice improved markers of placental efficiency and partially restored fetal growth. SFN supplementation was associated with reduced markers of fetal cardiac oxidative stress, including Nox2 and 3-nitrotyrosine, as well as attenuation of cardiac mass and cardiomyocyte area in male offspring by postnatal day 52 and improved vascular function in male and female offspring by postnatal day 98. Our findings are the first to highlight the functional consequences of NRF2 modulation in utero on early-life cardiovascular function in offspring, demonstrating that activation of NRF2 affords cardiovascular protection in offspring of pregnancies affected by redox dysregulation.
Keywords: Cardiovascular; Diabetes; NRF2; Obesity; Pregnancy; Sulforaphane.
Crown Copyright © 2023. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Coughlan M.T., Vervaart P.P., Permezel M., Georgiou H.M., Rice G.E. Altered placental oxidative stress status in gestational diabetes mellitus. Placenta. 2004;25(1):78–84. - PubMed
-
- Bugatto F., Quintero-Prado R., Visiedo F.M., Vilar-Sanchez J.M., Figueroa-Quinones A., Lopez-Tinoco C., Torrejon R., Bartha J.L. The influence of lipid and proinflammatory status on maternal uterine blood flow in women with late onset gestational diabetes. Reprod. Sci. 2018;25(6):837–843. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
