

Epigenomic dissection of Alzheimer's disease pinpoints causal variants and reveals epigenome erosion
- PMID: 37774680
- PMCID: PMC10782612
- DOI: 10.1016/j.cell.2023.08.040
Epigenomic dissection of Alzheimer's disease pinpoints causal variants and reveals epigenome erosion
Abstract
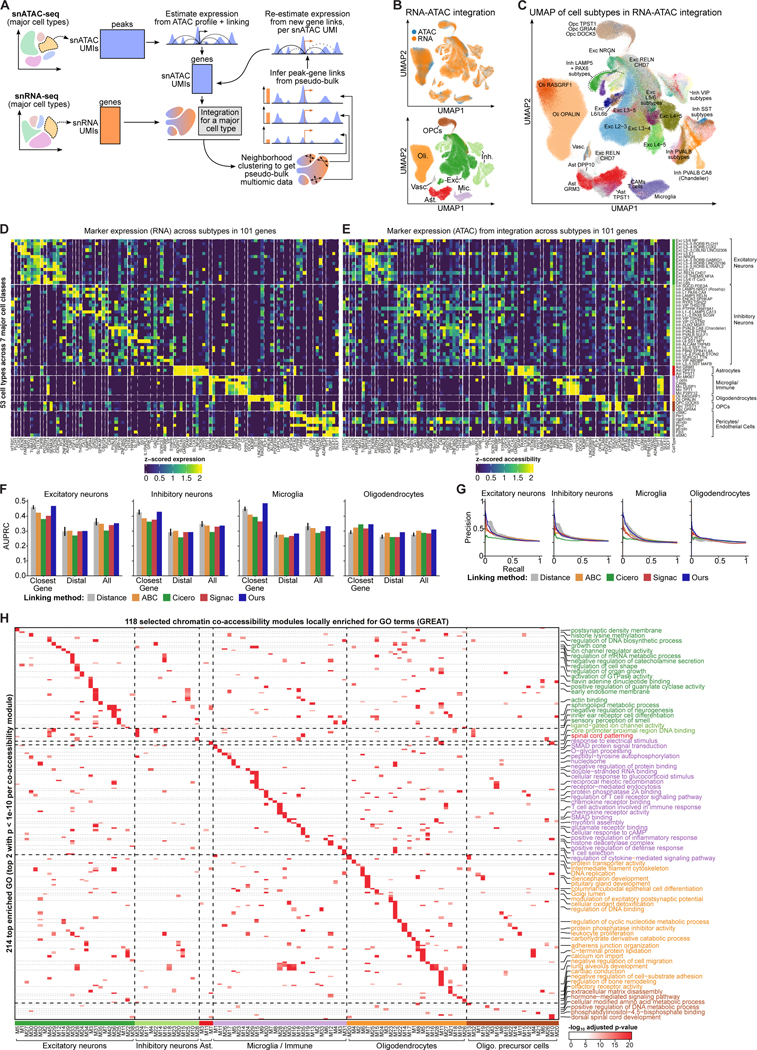
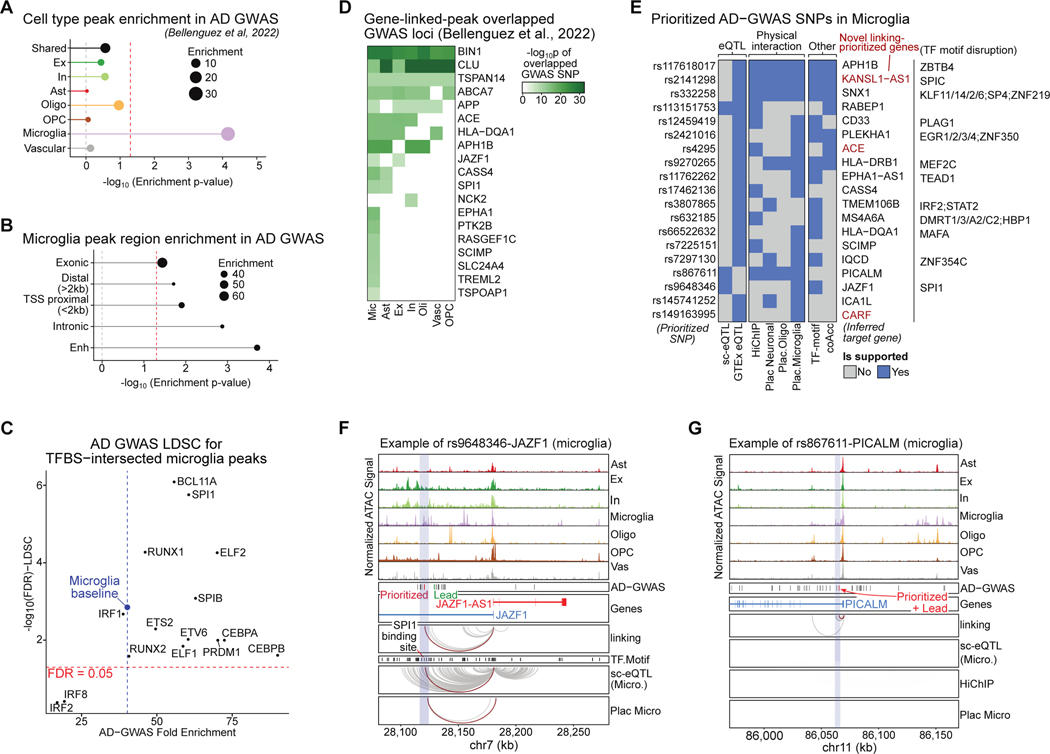
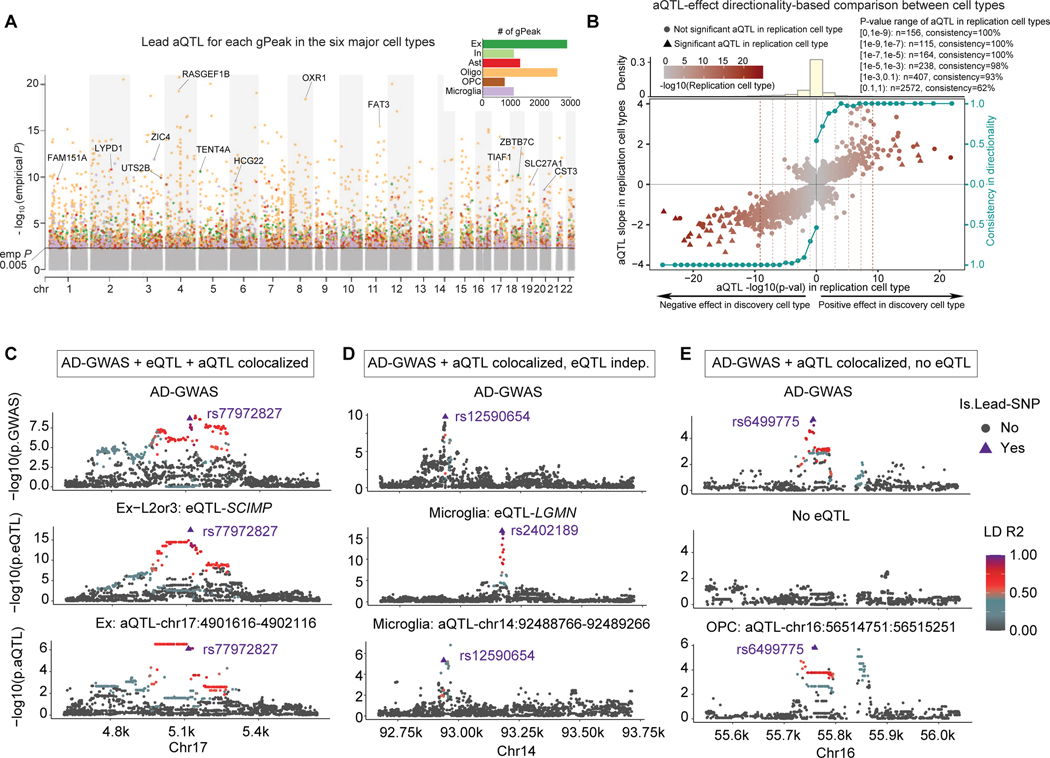
Recent work has identified dozens of non-coding loci for Alzheimer's disease (AD) risk, but their mechanisms and AD transcriptional regulatory circuitry are poorly understood. Here, we profile epigenomic and transcriptomic landscapes of 850,000 nuclei from prefrontal cortexes of 92 individuals with and without AD to build a map of the brain regulome, including epigenomic profiles, transcriptional regulators, co-accessibility modules, and peak-to-gene links in a cell-type-specific manner. We develop methods for multimodal integration and detecting regulatory modules using peak-to-gene linking. We show AD risk loci are enriched in microglial enhancers and for specific TFs including SPI1, ELF2, and RUNX1. We detect 9,628 cell-type-specific ATAC-QTL loci, which we integrate alongside peak-to-gene links to prioritize AD variant regulatory circuits. We report differential accessibility of regulatory modules in late AD in glia and in early AD in neurons. Strikingly, late-stage AD brains show global epigenome dysregulation indicative of epigenome erosion and cell identity loss.
Keywords: ATAC-QTL; Alzheimer’s disease; GWAS; epigenome; epigenome erosion; fine-mapping; multimodal integration; peak-to-gene linking.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests L.-H.T. is a member of the Scientific Advisory Boards of Cognito Therapeutics, 4M Therapeutics, Cell Signaling Technology, and Souvien Therapeutics, which have no association to the work described in this manuscript.
Figures



Comment in
-
The AD odyssey 2023: Tales of single cell.Cell. 2023 Sep 28;186(20):4257-4259. doi: 10.1016/j.cell.2023.09.001. Cell. 2023. PMID: 37774675
References
-
- Bennett DA, Wilson RS, Schneider JA, Evans DA, Beckett LA, Aggarwal NT, Barnes LL, Fox JH, and Bach J. (2002). Natural history of mild cognitive impairment in older persons. Neurology 59, 198–205. - PubMed
-
- Bennett DA, Schneider JA, Wilson RS, Bienias JL, and Arnold SE (2004). Neurofibrillary tangles mediate the association of amyloid load with clinical Alzheimer disease and level of cognitive function. Arch. Neurol 61, 378–384. - PubMed
-
- Braak H, and Braak E. (1995). Staging of Alzheimer’s disease-related neurofibrillary changes. Neurobiol. Aging 16, 271–278; discussion 278–284. - PubMed
-
- Querfurth HW, and LaFerla FM (2010). Alzheimer’s disease. N. Engl. J. Med 362, 329–344. - PubMed
Publication types
MeSH terms
Grants and funding
- T32 GM139790/GM/NIGMS NIH HHS/United States
- R01 AG074003/AG/NIA NIH HHS/United States
- RF1 AG062377/AG/NIA NIH HHS/United States
- P30 AG010161/AG/NIA NIH HHS/United States
- R01 AG062335/AG/NIA NIH HHS/United States
- U01 MH119509/MH/NIMH NIH HHS/United States
- R01 AG015819/AG/NIA NIH HHS/United States
- P30 AG072975/AG/NIA NIH HHS/United States
- U01 AG077227/AG/NIA NIH HHS/United States
- U01 DA053631/DA/NIDA NIH HHS/United States
- RF1 AG054012/AG/NIA NIH HHS/United States
- U01 AG046152/AG/NIA NIH HHS/United States
- R01 AG058002/AG/NIA NIH HHS/United States
- R01 HG008155/HG/NHGRI NIH HHS/United States
- U01 AG061356/AG/NIA NIH HHS/United States
- UG3 NS115064/NS/NINDS NIH HHS/United States
- R01 AG017917/AG/NIA NIH HHS/United States
- UH3 NS115064/NS/NINDS NIH HHS/United States
- R01 NS127187/NS/NINDS NIH HHS/United States
- RF1 NS129032/NS/NINDS NIH HHS/United States
- R01 AG081017/AG/NIA NIH HHS/United States
- RF1 AG057473/AG/NIA NIH HHS/United States
- U01 NS110453/NS/NINDS NIH HHS/United States
- R56 AG067151/AG/NIA NIH HHS/United States
- R01 AG015473/AG/NIA NIH HHS/United States
- R01 AG067151/AG/NIA NIH HHS/United States
- R01 MH109978/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
