

PET-measured human dopamine synthesis capacity and receptor availability predict trading rewards and time-costs during foraging
- PMID: 37777515
- PMCID: PMC10542376
- DOI: 10.1038/s41467-023-41897-0
PET-measured human dopamine synthesis capacity and receptor availability predict trading rewards and time-costs during foraging
Abstract
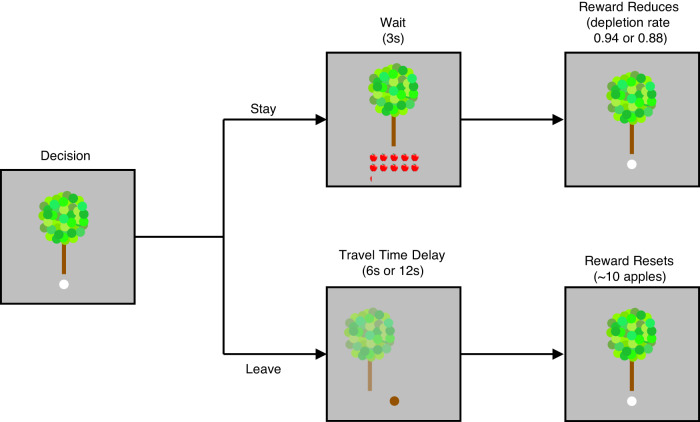
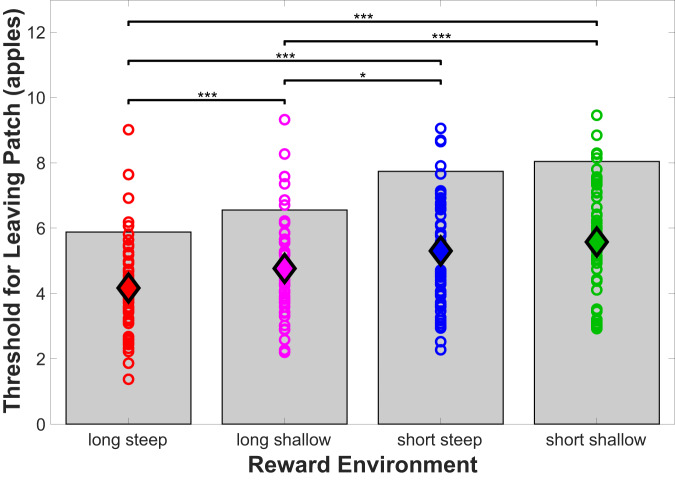
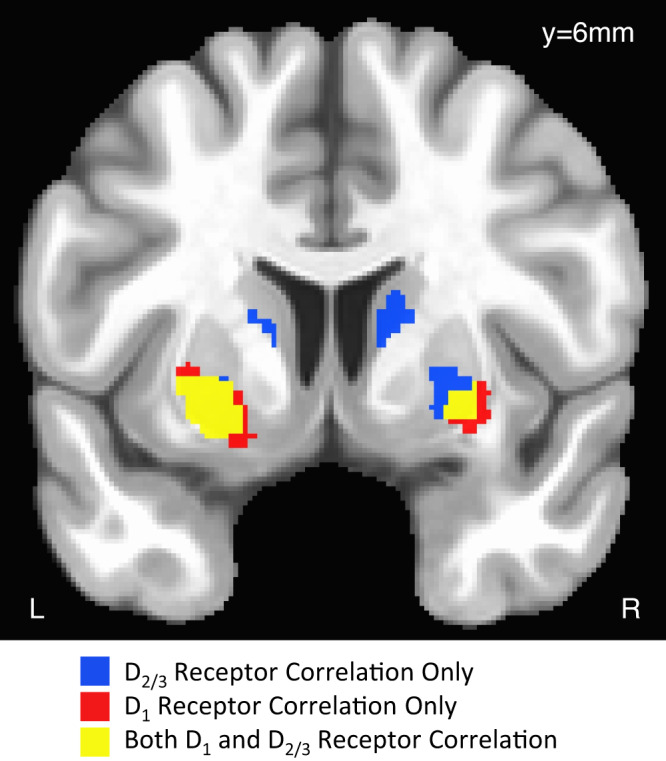
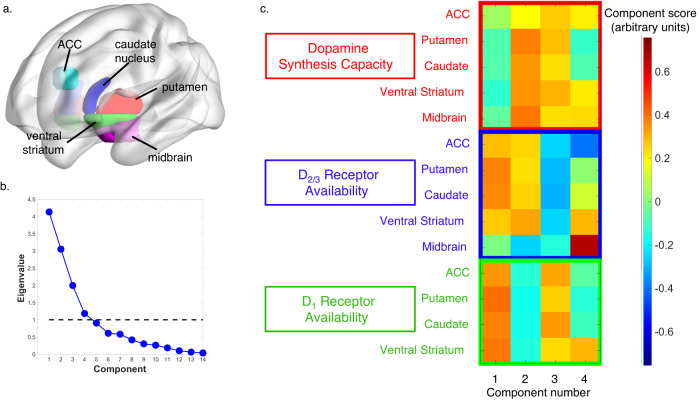
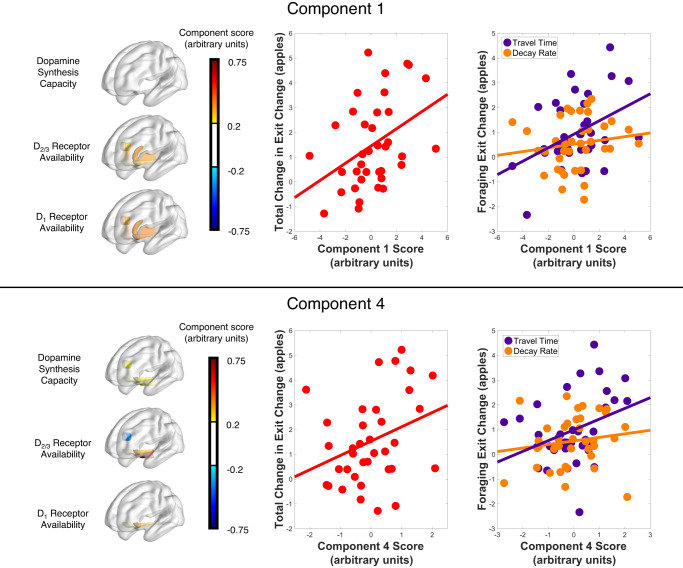
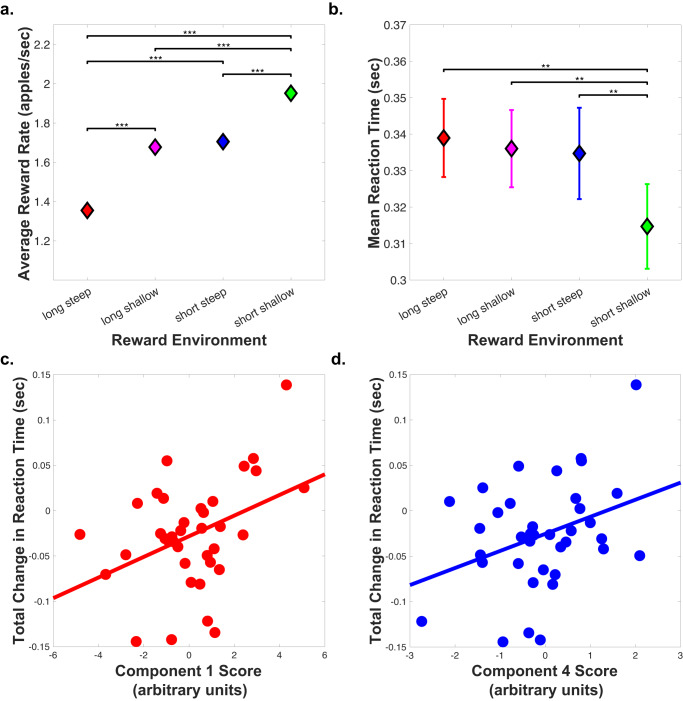
Foraging behavior requires weighing costs of time to decide when to leave one reward patch to search for another. Computational and animal studies suggest that striatal dopamine is key to this process; however, the specific role of dopamine in foraging behavior in humans is not well characterized. We use positron emission tomography (PET) imaging to directly measure dopamine synthesis capacity and D1 and D2/3 receptor availability in 57 healthy adults who complete a computerized foraging task. Using voxelwise data and principal component analysis to identify patterns of variation across PET measures, we show that striatal D1 and D2/3 receptor availability and a pattern of mesolimbic and anterior cingulate cortex dopamine function are important for adjusting the threshold for leaving a patch to explore, with specific sensitivity to changes in travel time. These findings suggest a key role for dopamine in trading reward benefits against temporal costs to modulate behavioral adaptions to changes in the reward environment critical for foraging.
Trial registration: ClinicalTrials.gov NCT00004571 NCT00942981 NCT00024622.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Charnov EL. Optimal foraging, the marginal value theorem. Theor. Popul. Biol. 1976;9:129–136. - PubMed
