

The roles of DGAT1 and DGAT2 in human myotubes are dependent on donor patho-physiological background
- PMID: 37779421
- PMCID: PMC10947296
- DOI: 10.1096/fj.202300960RR
The roles of DGAT1 and DGAT2 in human myotubes are dependent on donor patho-physiological background
Abstract
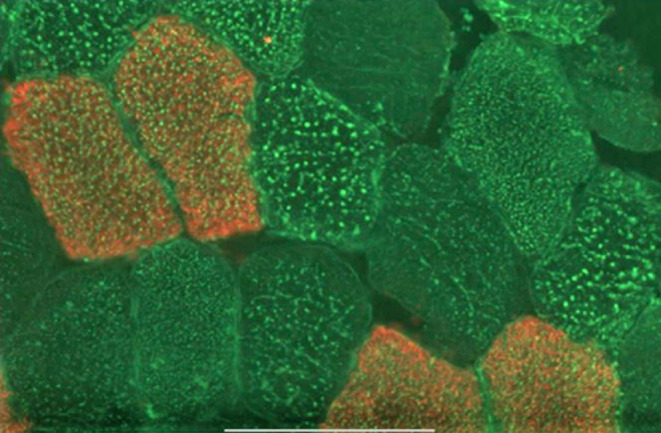
The roles of DGAT1 and DGAT2 in lipid metabolism and insulin responsiveness of human skeletal muscle were studied using cryosections and myotubes prepared from muscle biopsies from control, athlete, and impaired glucose regulation (IGR) cohorts of men. The previously observed increases in intramuscular triacylglycerol (IMTG) in athletes and IGR were shown to be related to an increase in lipid droplet (LD) area in type I fibers in athletes but, conversely, in type II fibers in IGR subjects. Specific inhibition of both diacylglycerol acyltransferase (DGAT) 1 and 2 decreased fatty acid (FA) uptake by myotubes, whereas only DGAT2 inhibition also decreased fatty acid oxidation. Fatty acid uptake in myotubes was negatively correlated with the lactate thresholds of the respective donors. DGAT2 inhibition lowered acetate uptake and oxidation in myotubes from all cohorts whereas DGAT1 inhibition had no effect. A positive correlation between acetate oxidation in myotubes and resting metabolic rate (RMR) from fatty acid oxidation in vivo was observed. Myotubes from athletes and IGR had higher rates of de novo lipogenesis from acetate that were normalized by DGAT2 inhibition. Moreover, DGAT2 inhibition in myotubes also resulted in increased insulin-induced Akt phosphorylation. The differential effects of DGAT1 and DGAT2 inhibition suggest that the specialized role of DGAT2 in esterifying nascent diacylglycerols and de novo synthesized FA is associated with synthesis of a pool of triacylglycerol, which upon hydrolysis results in effectors that promote mitochondrial fatty acid oxidation but decrease insulin signaling in skeletal muscle cells.
Keywords: diacylglycerol acyltransferases; fatty acid oxidation; insulin resistance; lipogenesis; muscle lipid; triacylglycerols.
© 2023 The Authors. The FASEB Journal published by Wiley Periodicals LLC on behalf of Federation of American Societies for Experimental Biology.
Figures








References
-
- Goodpaster BH, He J, Watkins S, Kelley DE. Skeletal muscle lipid content and insulin resistance: evidence for a paradox in endurance‐trained athletes. J Clin Endocrinol Metab. 2001;86:5755‐5761. - PubMed
-
- Russell AP. Lipotoxicity: the obese and endurance‐trained paradox. Int J Obes Relat Metab Disord. 2004;28(Suppl 4):S66‐S71. - PubMed
-
- Chavez JA, Summers SA. A ceramide‐centric view of insulin resistance. Cell Metab. 2012;15:585‐594. - PubMed
-
- Bergman BC, Goodpaster BH. Exercise and muscle lipid content, composition, and localization: influence on muscle insulin sensitivity. Diabetes. 2020;69:848‐858. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
