

p90RSK2, a new MLCK mediates contractility in myosin light chain kinase null smooth muscle
- PMID: 37781225
- PMCID: PMC10533999
- DOI: 10.3389/fphys.2023.1228488
p90RSK2, a new MLCK mediates contractility in myosin light chain kinase null smooth muscle
Abstract
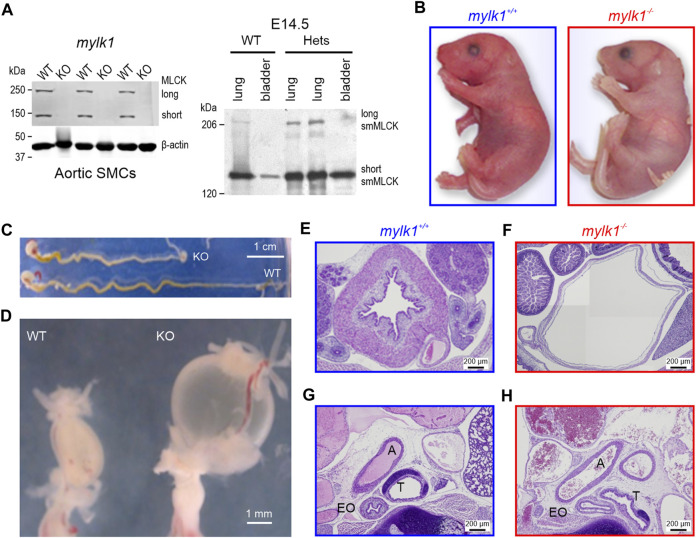
Introduction: Phosphorylation of smooth muscle (SM) myosin regulatory light chain (RLC20) is a critical switch leading to SM contraction. The canonical view held that only the short isoform of myosin light chain kinase (MLCK1) catalyzed this reaction. It is now accepted that auxiliary kinases may contribute to vascular SM tone and contractility. We have previously reported that p90 ribosomal S6 kinase (RSK2) functions as such a kinase, in parallel with MLCK1, contributing ∼25% of the maximal myogenic force in resistance arteries. Thus, RSK2 may be instrumental in the regulation of basal vascular tone and blood pressure. Here, we take advantage of a MLCK1 null mouse (mylk1 -/-) to further test our hypothesis that RSK2 can function as an MLCK, playing a significant physiological role in SM contractility. Methods: Using fetal (E14.5-18.5) SM tissues, as embryos die at birth, we investigated the necessity of MLCK for contractility and fetal development and determined the ability of RSK2 kinase to compensate for the lack of MLCK and characterized its signaling pathway in SM. Results and Discussion: Agonists induced contraction and RLC20 phosphorylation in mylk1 -/- SM was attenuated by RSK2 inhibition. The pCa-tension relationships in permeabilized strips of bladder showed no difference in Ca2+ sensitivity in WT vs mylk1 -/- muscles, although the magnitude of force responses was considerably smaller in the absence of MLCK. The magnitude of contractile responses was similar upon addition of GTPγS to activate the RhoA/ROCK pathway or calyculinA to inhibit the myosin phosphatase. The Ca2+-dependent tyrosine kinase, Pyk2, contributed to RSK2-mediated contractility and RLC20 phosphorylation. Proximity-ligation and immunoprecipitation assays demonstrated an association of RSK2, PDK1 and ERK1/2 with MLCK and actin. RSK2, PDK1, ERK1/2 and MLCK formed a signaling complex on the actin filament, positioning them for interaction with adjacent myosin heads. The Ca2+-dependent component reflected the agonist mediated increases in Ca2+, which activated the Pyk2/PDK1/RSK2 signaling cascade. The Ca2+-independent component was through activation of Erk1/2/PDK1/RSK2 leading to direct phosphorylation of RLC20, to increase contraction. Overall, RSK2 signaling constitutes a new third signaling pathway, in addition to the established Ca2+/CaM/MLCK and RhoA/ROCK pathways to regulate SM contractility.
Keywords: MLCK; PDK1; Pyk2; RSK2; mylk1; p90 ribosomal S6 kinase; smooth muscle.
Copyright © 2023 Kalra, Artamonov, Wang, Franke, Markowska, Jin, Derewenda, Ayon and Somlyo.
Conflict of interest statement
Author AF was employed by Brain Surgery Worldwide. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





Update of
-
p90RSK2, a new MLCK, rescues contractility in myosin light chain kinase null smooth muscle.bioRxiv [Preprint]. 2023 May 22:2023.05.22.541840. doi: 10.1101/2023.05.22.541840. bioRxiv. 2023. Update in: Front Physiol. 2023 Sep 13;14:1228488. doi: 10.3389/fphys.2023.1228488. PMID: 37292593 Free PMC article. Updated. Preprint.
References
-
- Artamonov M. V., Momotani K., Stevenson A., Trentham D. R., Derewenda U., Derewenda Z. S., et al. (2013b). Agonist-induced Ca2+ sensitization in smooth muscle: redundancy of rho guanine nucleotide exchange factors (RhoGEFs) and response kinetics, a caged compound study. J. Biol. Chem. 288, 34030–34040. [pii]. 10.1074/jbc.M113.514596 - DOI - PMC - PubMed
-
- Artamonov M. V., Sonkusare S. K., Good M. E., Momotani K., Eto M., Isakson B. E., et al. (2018). RSK2 contributes to myogenic vasoconstriction of resistance arteries by activating smooth muscle myosin and the Na(+)/H(+) exchanger. Sci. Signal 11, eaar3924. 10.1126/scisignal.aar3924 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous