

This is a preprint.
Immune memory shapes human polyclonal antibody responses to H2N2 vaccination
- PMID: 37781590
- PMCID: PMC10541104
- DOI: 10.1101/2023.08.23.554525
Immune memory shapes human polyclonal antibody responses to H2N2 vaccination
Update in
-
Immune memory shapes human polyclonal antibody responses to H2N2 vaccination.Cell Rep. 2024 May 28;43(5):114171. doi: 10.1016/j.celrep.2024.114171. Epub 2024 May 7. Cell Rep. 2024. PMID: 38717904 Free PMC article.
Abstract
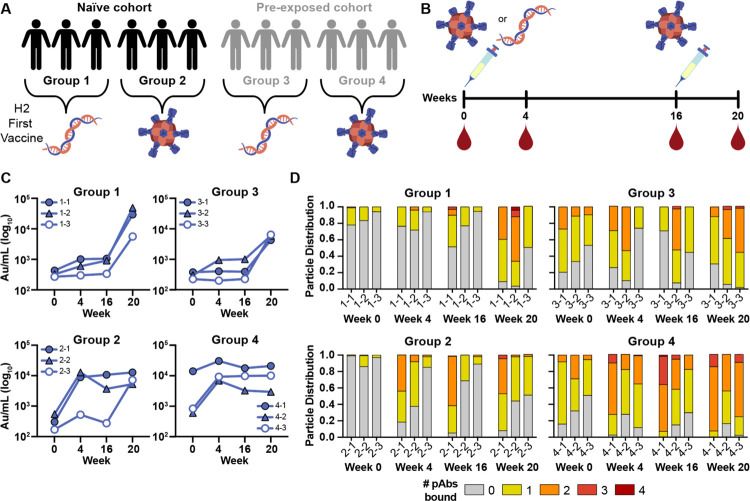
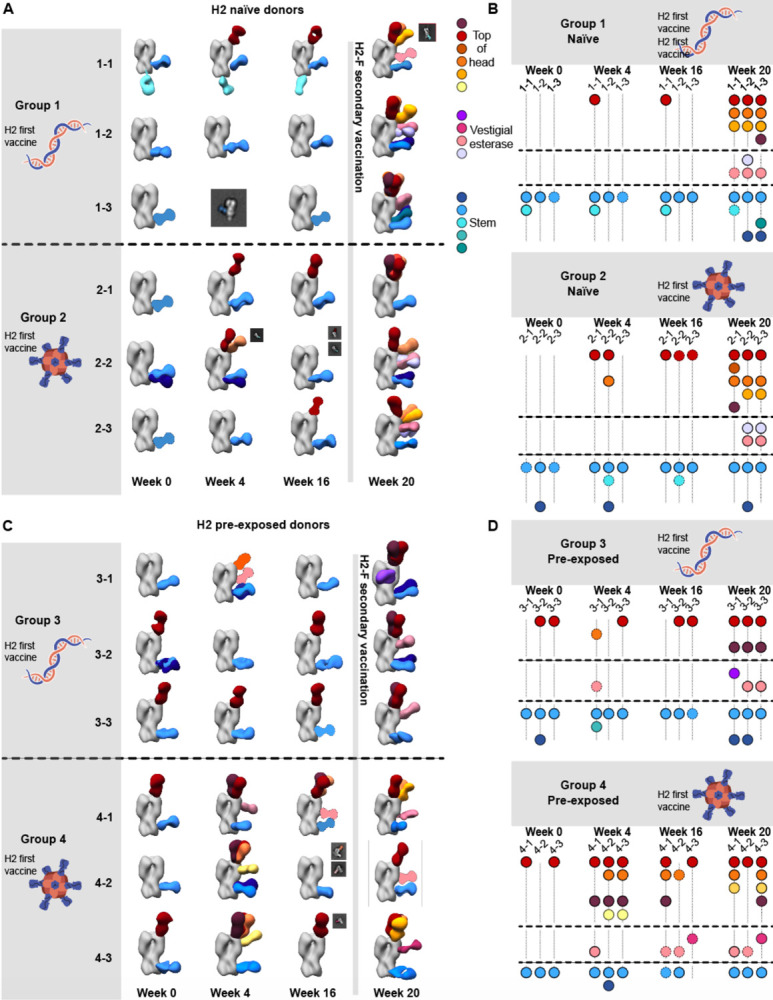
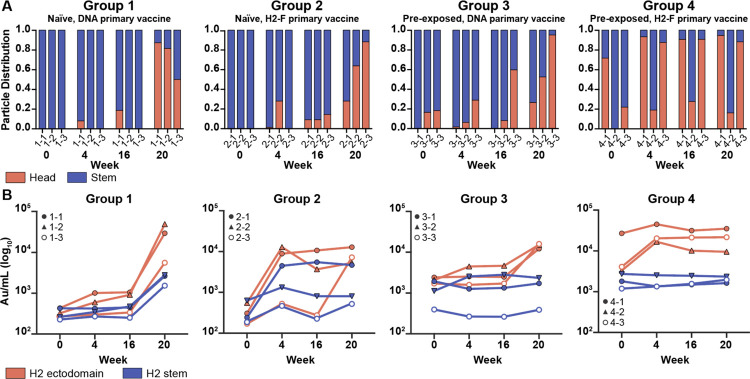
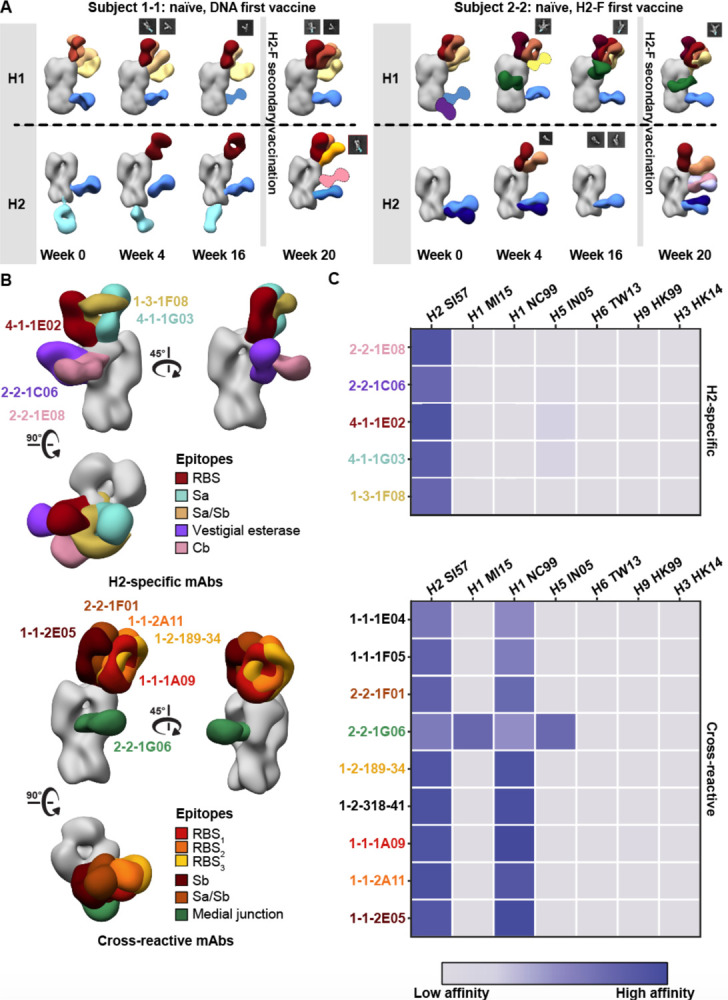
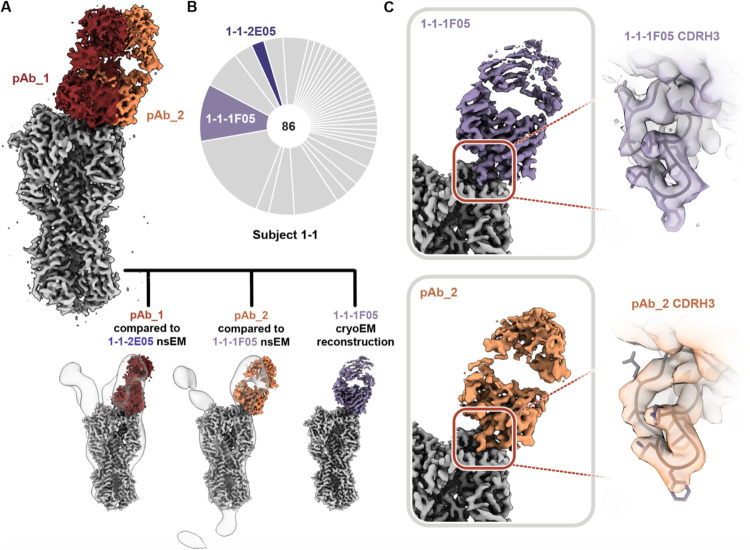
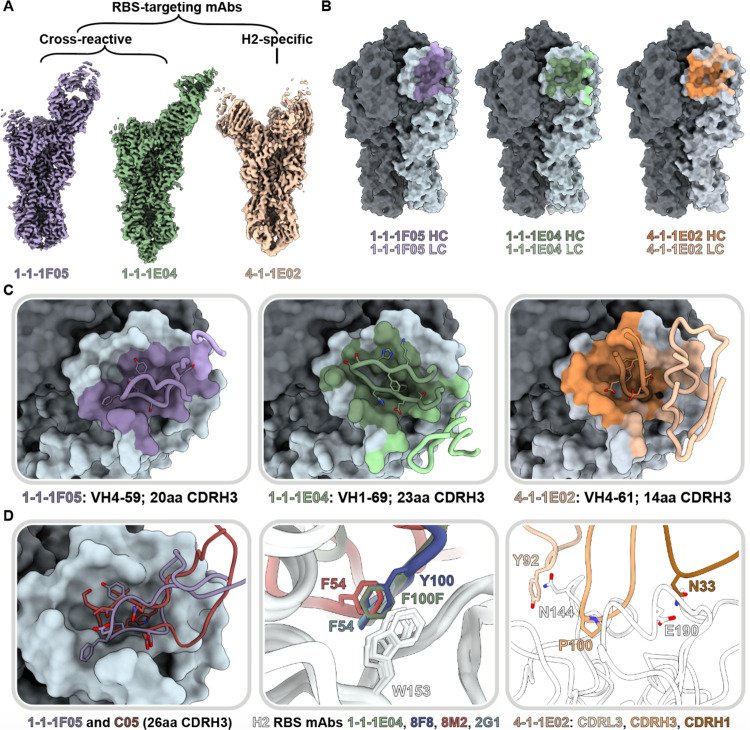
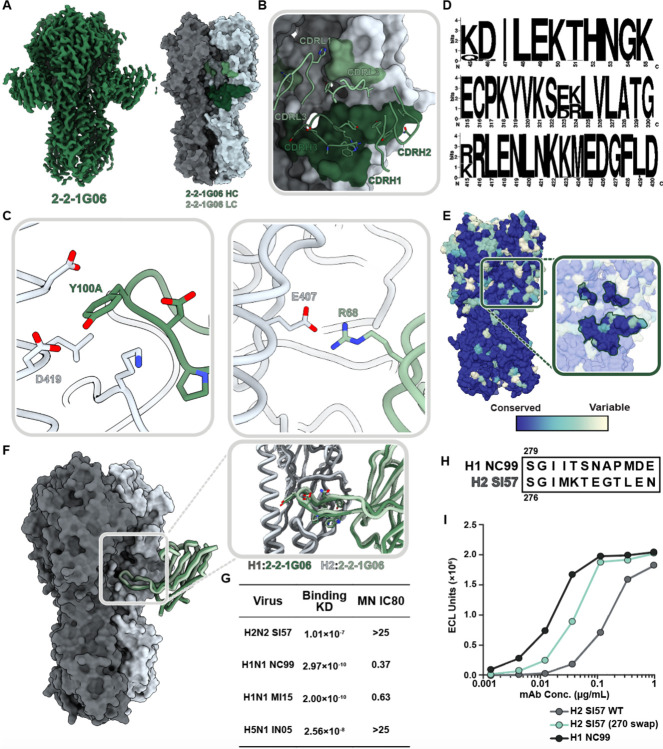
Influenza A virus subtype H2N2, which caused the 1957 influenza pandemic, remains a global threat. A recent phase I clinical trial investigating a ferritin nanoparticle displaying H2 hemagglutinin in H2-naïve and H2-exposed adults. Therefore, we could perform comprehensive structural and biochemical characterization of immune memory on the breadth and diversity of the polyclonal serum antibody response elicited after H2 vaccination. We temporally map the epitopes targeted by serum antibodies after first and second vaccinations and show previous H2 exposure results in higher responses to the variable head domain of hemagglutinin while initial responses in H2-naïve participants are dominated by antibodies targeting conserved epitopes. We use cryo-EM and monoclonal B cell isolation to describe the molecular details of cross-reactive antibodies targeting conserved epitopes on the hemagglutinin head including the receptor binding site and a new site of vulnerability deemed the medial junction. Our findings accentuate the impact of pre-existing influenza exposure on serum antibody responses.
Keywords: cryoEM; hemagglutinin; influenza; neutralizing antibody; structure-based vaccine design.
Conflict of interest statement
DECLARATION OF INTERESTS. The authors declare no conflicting interests.
Figures
References
-
- Cox N. J. & Subbarao K. GLOBAL EPIDEMIOLOGY OF INFLUENZA: Past and Present*. Annu. Rev. Med vol. 51 www.annualreviews.org (2000). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources