

This is a preprint.
Mapping Cellular Interactions from Spatially Resolved Transcriptomics Data
- PMID: 37781617
- PMCID: PMC10541142
- DOI: 10.1101/2023.09.18.558298
Mapping Cellular Interactions from Spatially Resolved Transcriptomics Data
Update in
-
Mapping cellular interactions from spatially resolved transcriptomics data.Nat Methods. 2024 Oct;21(10):1830-1842. doi: 10.1038/s41592-024-02408-1. Epub 2024 Sep 3. Nat Methods. 2024. PMID: 39227721
Abstract
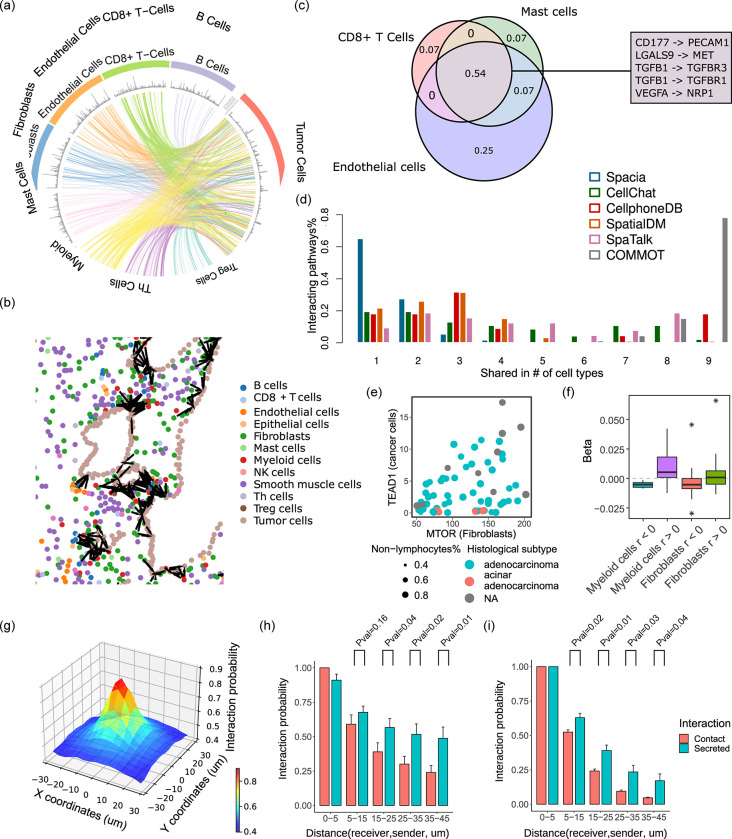
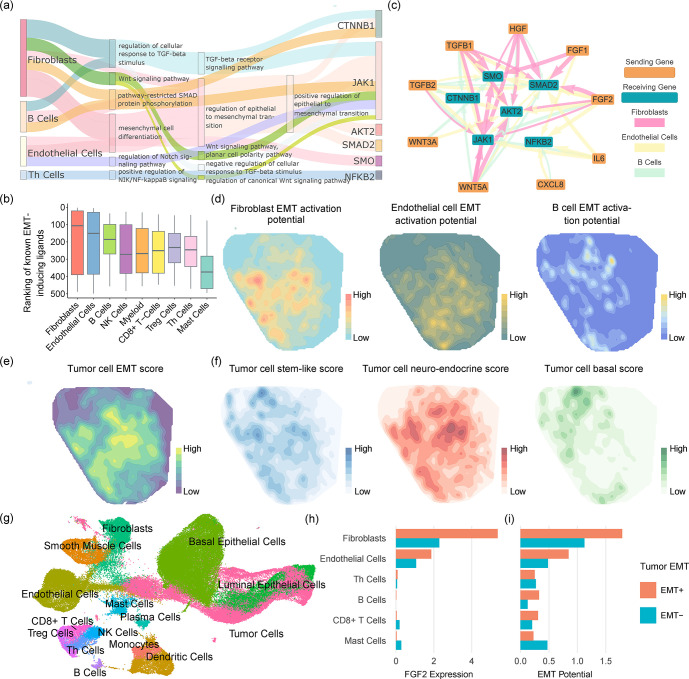
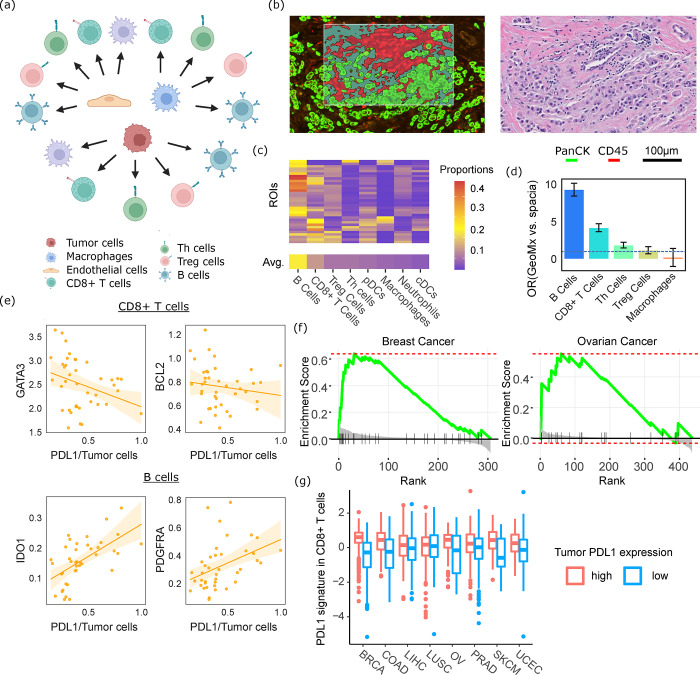
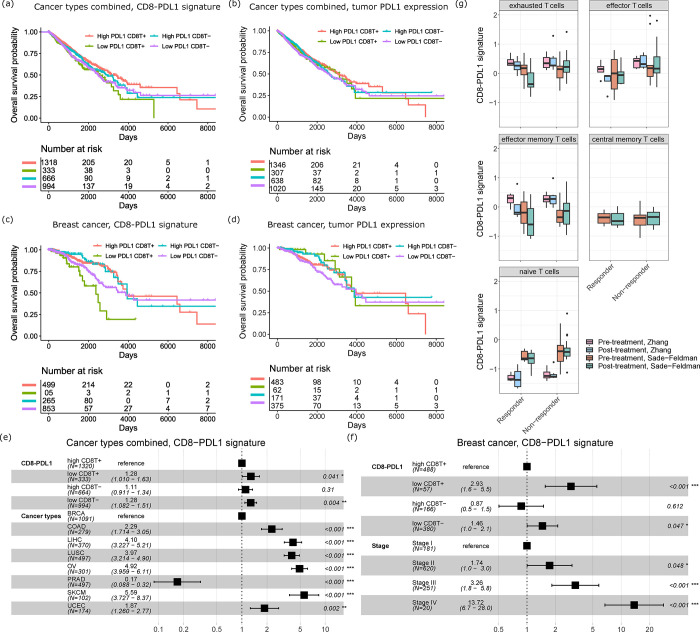
Cell-cell communication (CCC) is essential to how life forms and functions. However, accurate, high-throughput mapping of how expression of all genes in one cell affects expression of all genes in another cell is made possible only recently, through the introduction of spatially resolved transcriptomics technologies (SRTs), especially those that achieve single cell resolution. However, significant challenges remain to analyze such highly complex data properly. Here, we introduce a Bayesian multi-instance learning framework, spacia, to detect CCCs from data generated by SRTs, by uniquely exploiting their spatial modality. We highlight spacia's power to overcome fundamental limitations of popular analytical tools for inference of CCCs, including losing single-cell resolution, limited to ligand-receptor relationships and prior interaction databases, high false positive rates, and most importantly the lack of consideration of the multiple-sender-to-one-receiver paradigm. We evaluated the fitness of spacia for all three commercialized single cell resolution ST technologies: MERSCOPE/Vizgen, CosMx/Nanostring, and Xenium/10X. Spacia unveiled how endothelial cells, fibroblasts and B cells in the tumor microenvironment contribute to Epithelial-Mesenchymal Transition and lineage plasticity in prostate cancer cells. We deployed spacia in a set of pan-cancer datasets and showed that B cells also participate in PDL1/PD1 signaling in tumors. We demonstrated that a CD8+ T cell/PDL1 effectiveness signature derived from spacia analyses is associated with patient survival and response to immune checkpoint inhibitor treatments in 3,354 patients. We revealed differential spatial interaction patterns between γδ T cells and liver hepatocytes in healthy and cancerous contexts. Overall, spacia represents a notable step in advancing quantitative theories of cellular communications.
Keywords: GeoMX; MERSCOPE; cellular interaction; spacia; spatially resolved transcriptomics.
Conflict of interest statement
COMPETING INTEREST STATEMENT Tao Wang is one of the scientific co-founders of NightStar Biotechnologies, Inc. Ariella Hanker receives or has received research grants from Takeda and Lilly and nonfinancial support from Puma Biotechnology and Tempus. Carlos Arteaga receives or has received research grants from Pfizer, Lilly, and Takeda; holds minor stock options in Provista; serves or has served in an advisory role to Novartis, Merck, Lilly, Daiichi Sankyo, Taiho Oncology, OrigiMed, Puma Biotechnology, Immunomedics, AstraZeneca, Arvinas, and Sanofi; and reports scientific advisory board remuneration from the Susan G. Komen Foundation.
Figures

References
-
- Bechtel T. J., Reyes-Robles T., Fadeyi O. O. & Oslund R. C. Strategies for monitoring cell-cell interactions. Nat. Chem. Biol. 17, 641–652 (2021). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials