

Mice lacking triglyceride synthesis enzymes in adipose tissue are resistant to diet-induced obesity
- PMID: 37782317
- PMCID: PMC10545428
- DOI: 10.7554/eLife.88049
Mice lacking triglyceride synthesis enzymes in adipose tissue are resistant to diet-induced obesity
Abstract
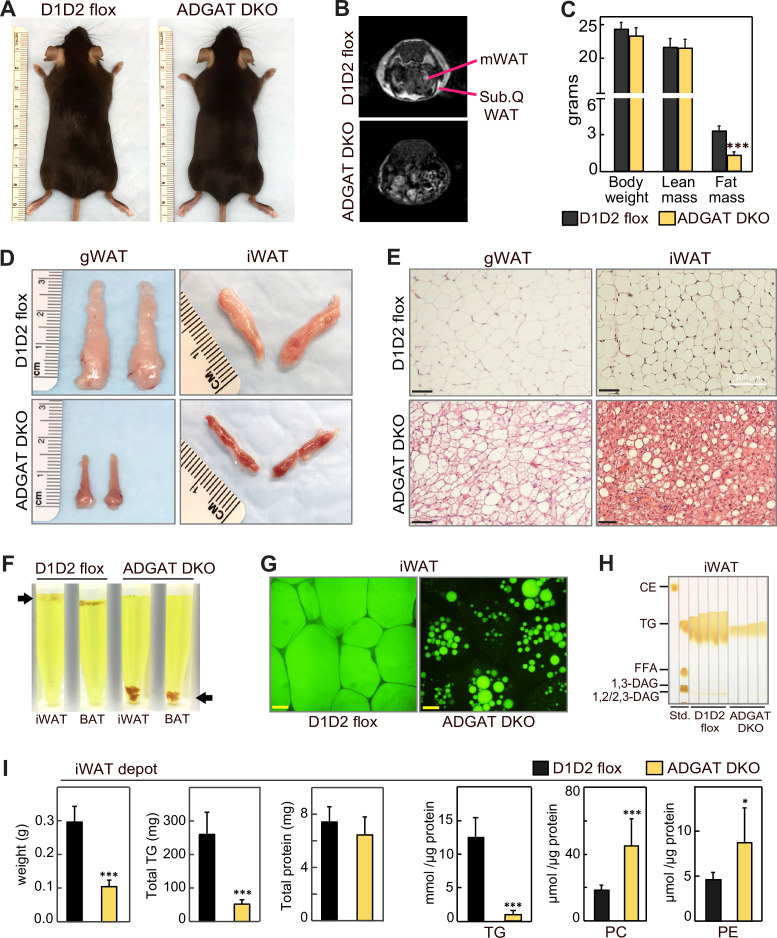
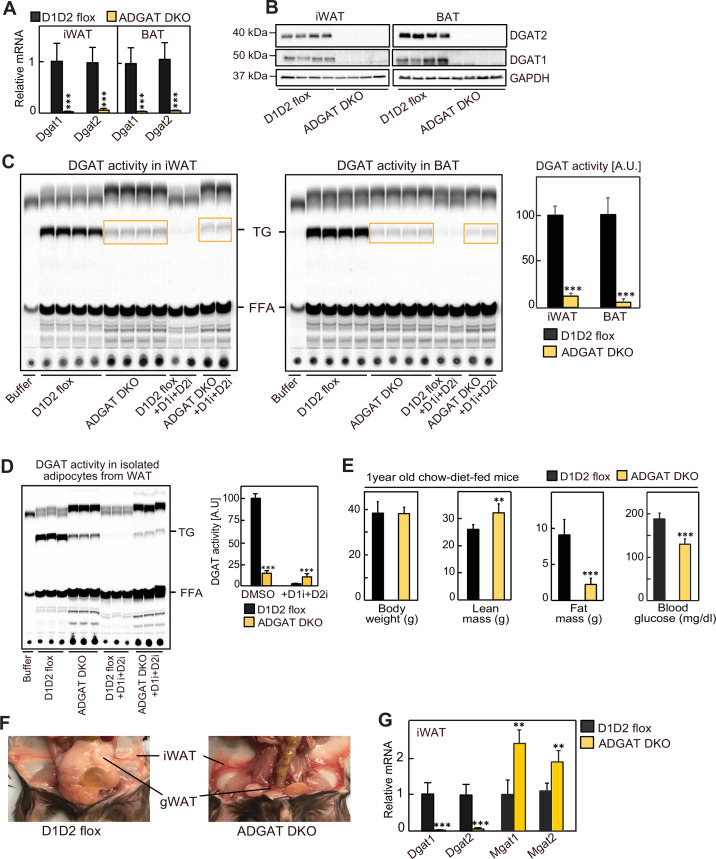
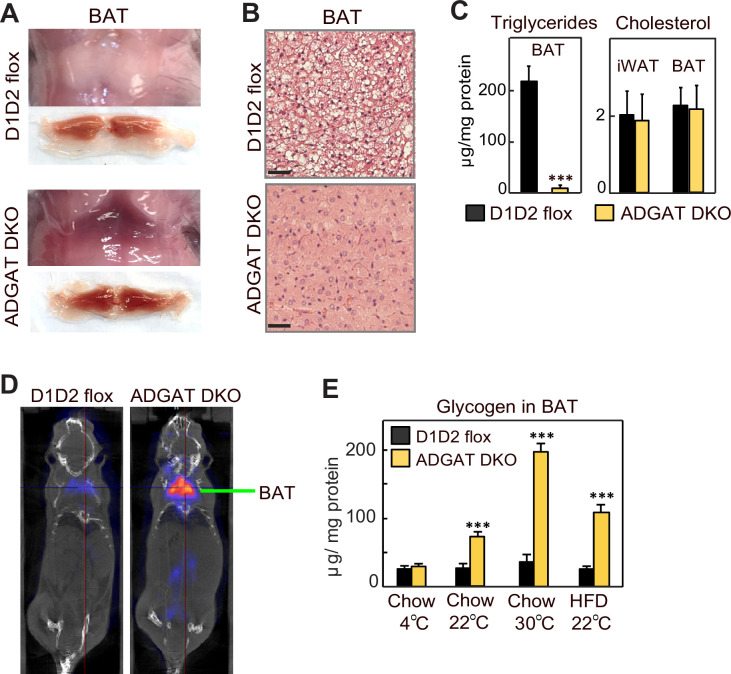
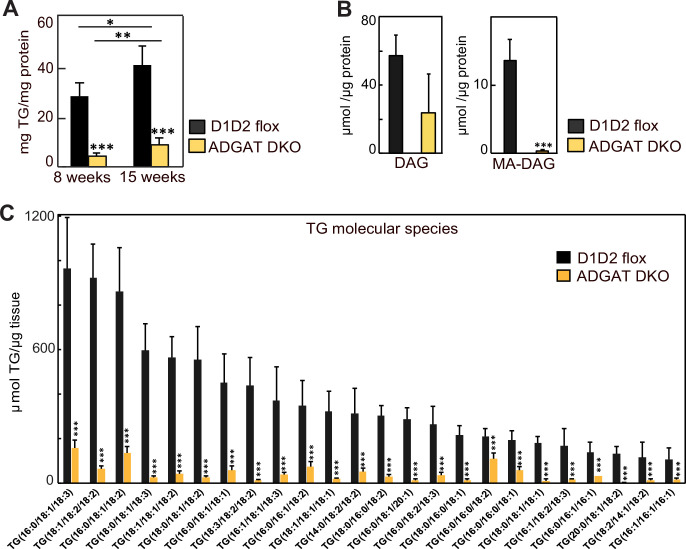
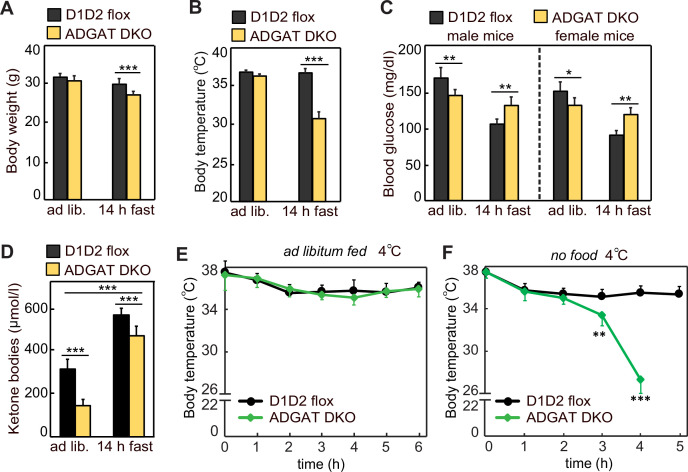
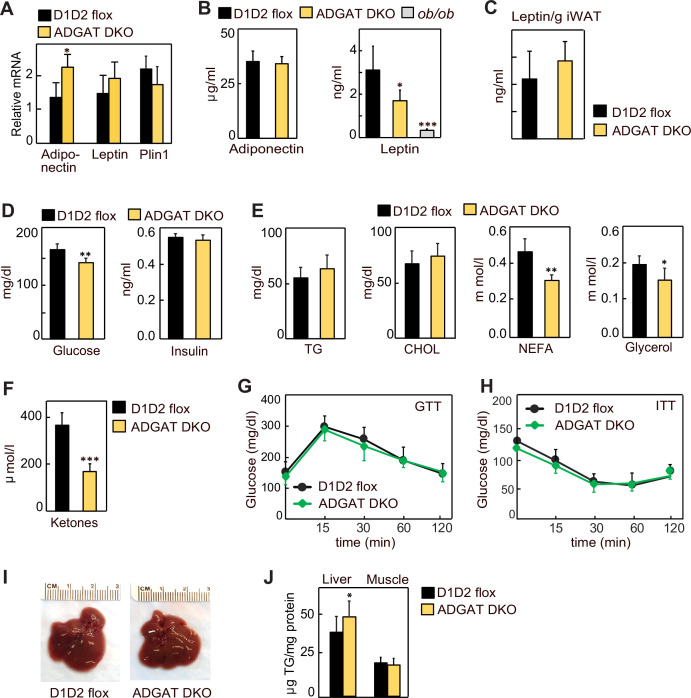
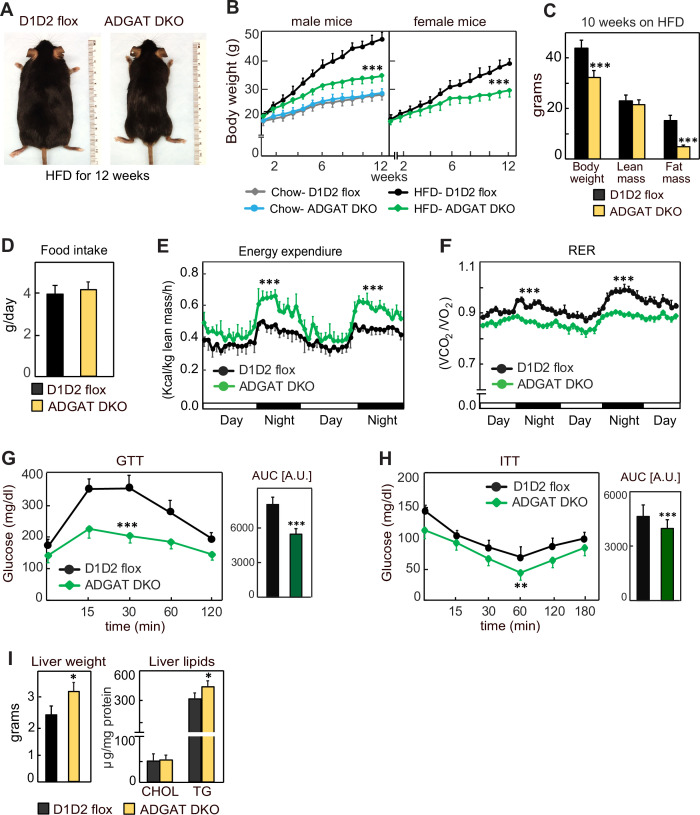
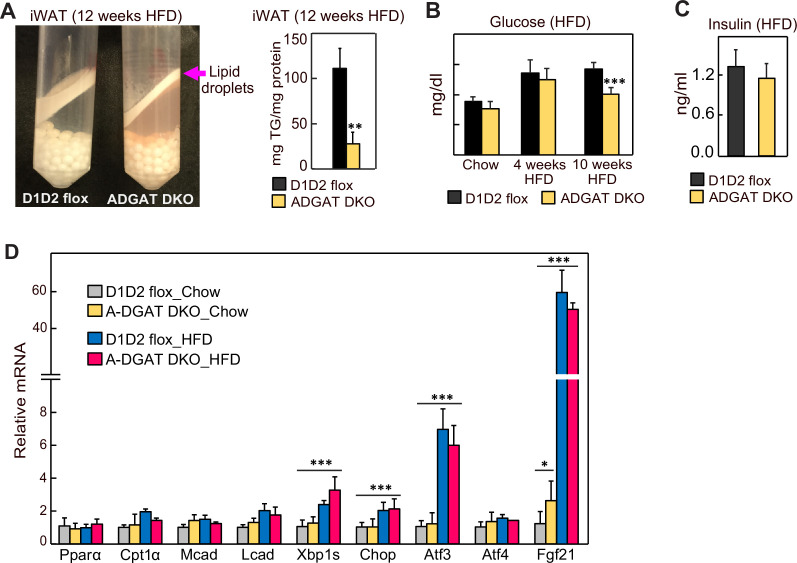
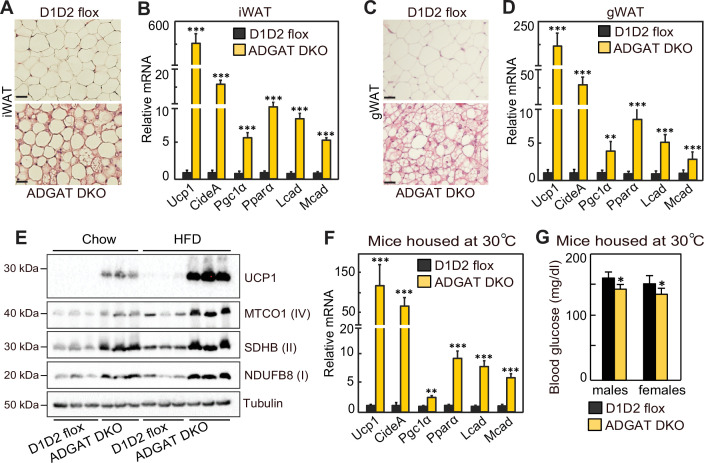
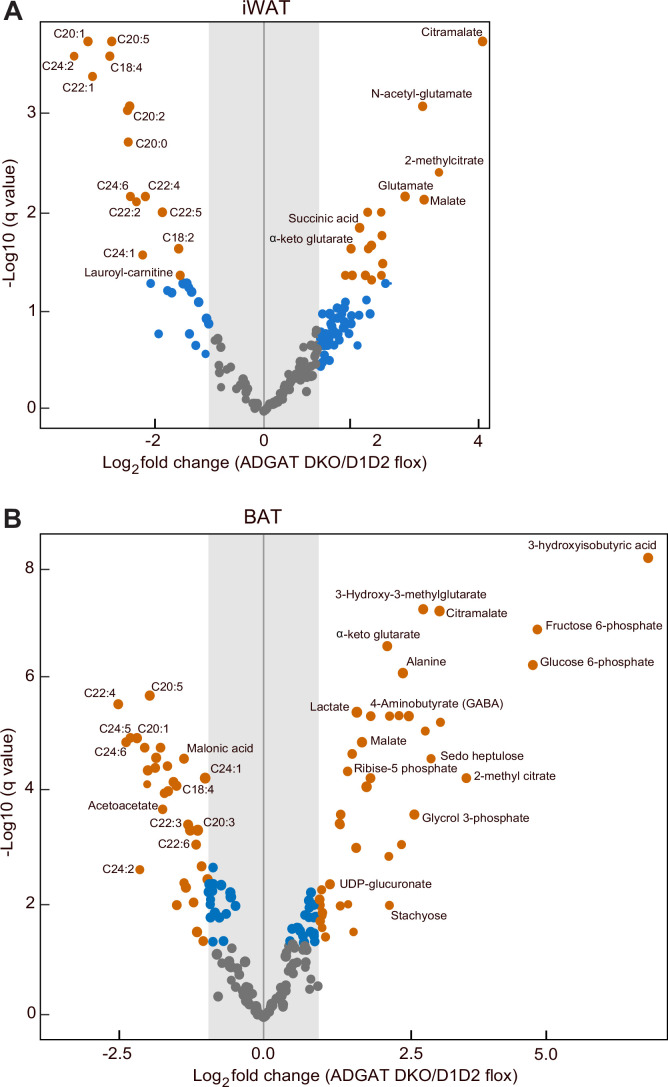
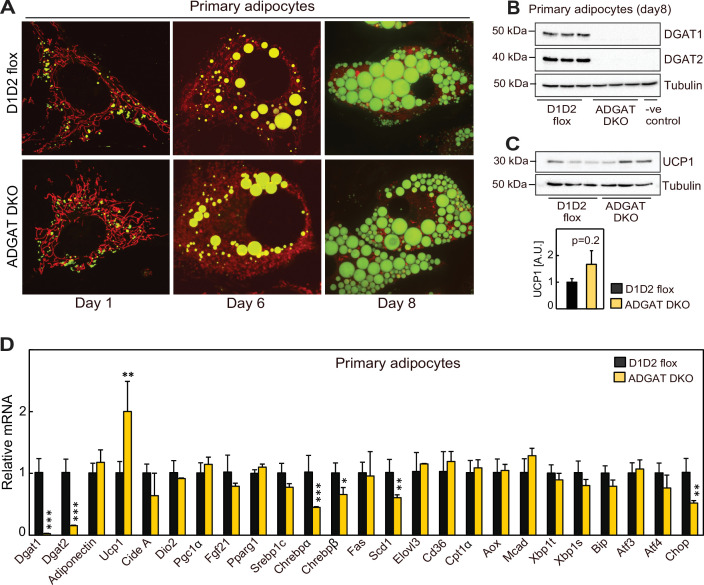
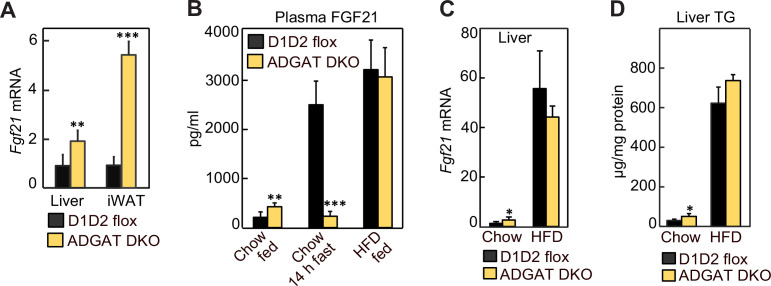
Triglycerides (TGs) in adipocytes provide the major stores of metabolic energy in the body. Optimal amounts of TG stores are desirable as insufficient capacity to store TG, as in lipodystrophy, or exceeding the capacity for storage, as in obesity, results in metabolic disease. We hypothesized that mice lacking TG storage in adipocytes would result in excess TG storage in cell types other than adipocytes and severe lipotoxicity accompanied by metabolic disease. To test this hypothesis, we selectively deleted both TG synthesis enzymes, DGAT1 and DGAT2, in adipocytes (ADGAT DKO mice). As expected with depleted energy stores, ADGAT DKO mice did not tolerate fasting well and, with prolonged fasting, entered torpor. However, ADGAT DKO mice were unexpectedly otherwise metabolically healthy and did not accumulate TGs ectopically or develop associated metabolic perturbations, even when fed a high-fat diet. The favorable metabolic phenotype resulted from activation of energy expenditure, in part via BAT (brown adipose tissue) activation and beiging of white adipose tissue. Thus, the ADGAT DKO mice provide a fascinating new model to study the coupling of metabolic energy storage to energy expenditure.
Keywords: adipose tissue; fat; glucose metabolism; lipodystrophy; medicine; metabolism; mouse; triglyceride.
© 2023, Chitraju et al.
Conflict of interest statement
CC, AF, YA, KW, BY, SH, TW, RF No competing interests declared
Figures
Update of
- doi: 10.1101/2022.05.05.490833
- doi: 10.7554/eLife.88049.1
- doi: 10.7554/eLife.88049.2
References
-
- Abu-Odeh M, Zhang Y, Reilly SM, Ebadat N, Keinan O, Valentine JM, Hafezi-Bakhtiari M, Ashayer H, Mamoun L, Zhou X, Zhang J, Yu RT, Dai Y, Liddle C, Downes M, Evans RM, Kliewer SA, Mangelsdorf DJ, Saltiel AR. FGF21 promotes thermogenic gene expression as an autocrine factor in adipocytes. Cell Reports. 2021;35:109331. doi: 10.1016/j.celrep.2021.109331. - DOI - PMC - PubMed
-
- Cases S, Smith SJ, Zheng YW, Myers HM, Lear SR, Sande E, Novak S, Collins C, Welch CB, Lusis AJ, Erickson SK, Farese RV. Identification of a gene encoding an acyl CoA:diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis. PNAS. 1998;95:13018–13023. doi: 10.1073/pnas.95.22.13018. - DOI - PMC - PubMed
-
- Chen HC, Smith SJ, Ladha Z, Jensen DR, Ferreira LD, Pulawa LK, McGuire JG, Pitas RE, Eckel RH, Farese RV. Increased insulin and leptin sensitivity in mice lacking acyl CoA:diacylglycerol acyltransferase 1. The Journal of Clinical Investigation. 2002;109:1049–1055. doi: 10.1172/JCI14672. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
