

VP2 residue N142 of coxsackievirus A10 is critical for the interaction with KREMEN1 receptor and neutralizing antibodies and the pathogenicity in mice
- PMID: 37788227
- PMCID: PMC10547193
- DOI: 10.1371/journal.ppat.1011662
VP2 residue N142 of coxsackievirus A10 is critical for the interaction with KREMEN1 receptor and neutralizing antibodies and the pathogenicity in mice
Abstract
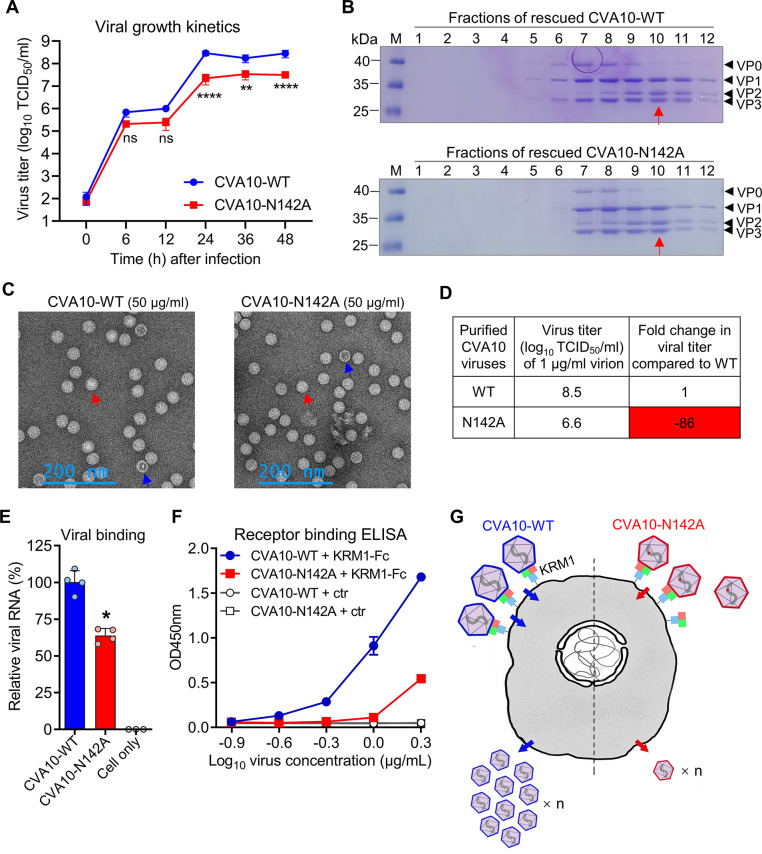
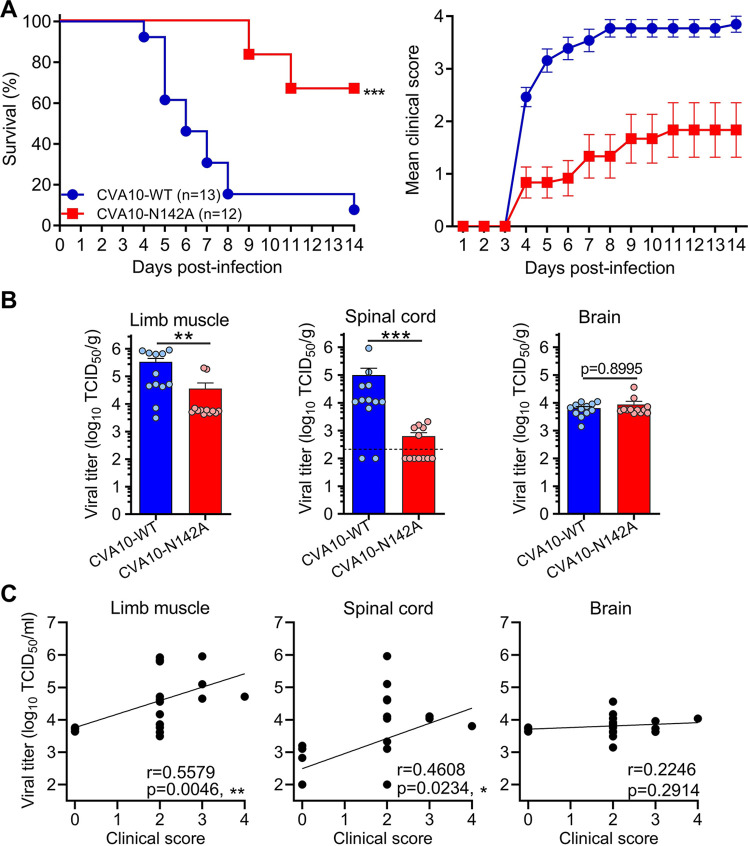
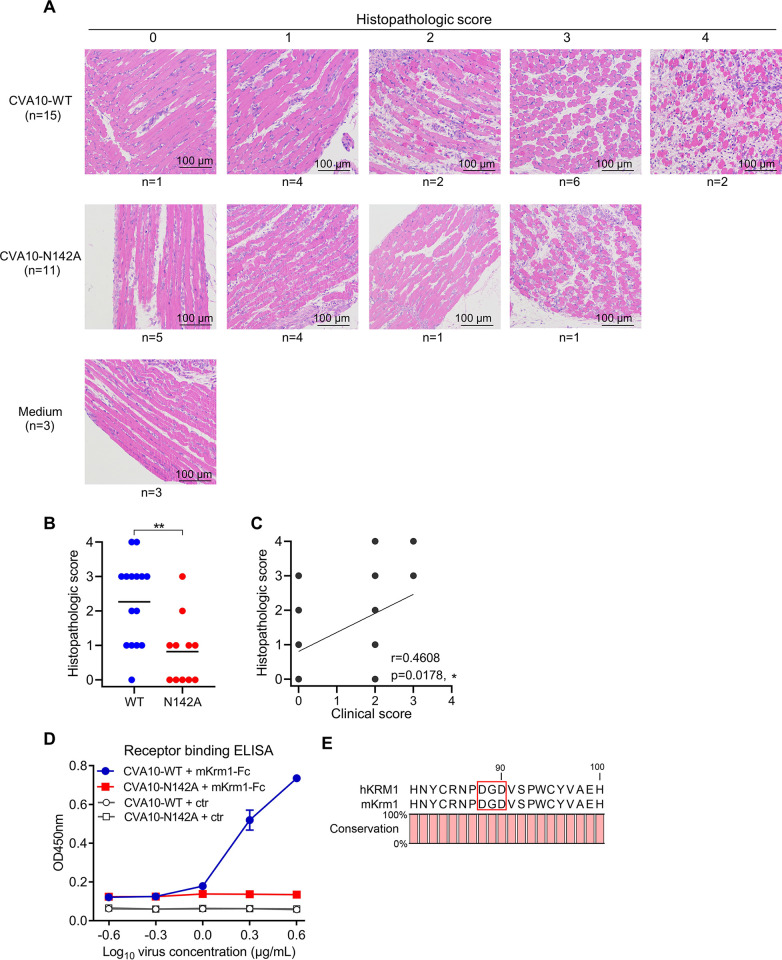
Coxsackievirus A10 (CVA10) has recently emerged as one of the major causative agents of hand, foot, and mouth disease. CVA10 may also cause a variety of complications. No approved vaccine or drug is currently available for CVA10. The residues of CVA10 critical for viral attachment, infectivity and in vivo pathogenicity have not been identified by experiment. Here, we report the identification of CVA10 residues important for binding to cellular receptor KREMEN1. We identified VP2 N142 as a key receptor-binding residue by screening of CVA10 mutants resistant to neutralization by soluble KREMEN1 protein. The receptor-binding residue N142 is exposed on the canyon rim but highly conserved in all naturally occurring CVA10 strains, which provides a counterexample to the canyon hypothesis. Residue N142 when mutated drastically reduced receptor-binding activity, resulting in decreased viral attachment and infection in cell culture. More importantly, residue N142 when mutated reduced viral replication in limb muscle and spinal cord of infected mice, leading to lower mortality and less severe clinical symptoms. Additionally, residue N142 when mutated could decrease viral binding affinity to anti-CVA10 polyclonal antibodies and a neutralizing monoclonal antibody and render CVA10 resistant to neutralization by the anti-CVA10 antibodies. Overall, our study highlights the essential role of VP2 residue N142 of CVA10 in the interactions with KREMEN1 receptor and neutralizing antibodies and viral virulence in mice, facilitating the understanding of the molecular mechanisms of CVA10 infection and immunity. Our study also provides important information for rational development of antibody-based treatment and vaccines against CVA10 infection.
Copyright: © 2023 Li et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Completely conserved VP2 residue K140 of KREMEN1-dependent enteroviruses is critical for virus-receptor interactions and viral infection.mBio. 2025 Feb 5;16(2):e0304024. doi: 10.1128/mbio.03040-24. Epub 2025 Jan 16. mBio. 2025. PMID: 39817751 Free PMC article.
-
Recombinant virus-like particle presenting a newly identified coxsackievirus A10 neutralization epitope induces protective immunity in mice.Antiviral Res. 2019 Apr;164:139-146. doi: 10.1016/j.antiviral.2019.02.016. Epub 2019 Feb 25. Antiviral Res. 2019. PMID: 30817941
-
Protective Efficacies of Formaldehyde-Inactivated Whole-Virus Vaccine and Antivirals in a Murine Model of Coxsackievirus A10 Infection.J Virol. 2017 Jun 9;91(13):e00333-17. doi: 10.1128/JVI.00333-17. Print 2017 Jul 1. J Virol. 2017. PMID: 28424287 Free PMC article.
-
Prospect and challenges for the development of multivalent vaccines against hand, foot and mouth diseases.Vaccine. 2014 Oct 29;32(47):6177-82. doi: 10.1016/j.vaccine.2014.08.064. Epub 2014 Sep 13. Vaccine. 2014. PMID: 25218294 Review.
-
Recent development of enterovirus A vaccine candidates for the prevention of hand, foot, and mouth disease.Expert Rev Vaccines. 2018 Sep;17(9):819-831. doi: 10.1080/14760584.2018.1510326. Epub 2018 Aug 22. Expert Rev Vaccines. 2018. PMID: 30095317 Review.
Cited by
-
Transition of D3c branch and novel recombination events contribute to the diversity of Coxsackievirus A6 in Beijing, China, from 2019 to 2023.Virus Evol. 2025 May 11;11(1):veaf036. doi: 10.1093/ve/veaf036. eCollection 2025. Virus Evol. 2025. PMID: 40574752 Free PMC article.
-
Identification of four neutralizing antigenic sites on the enterovirus D68 capsid.J Virol. 2023 Dec 21;97(12):e0160023. doi: 10.1128/jvi.01600-23. Epub 2023 Dec 4. J Virol. 2023. PMID: 38047678 Free PMC article.
-
Completely conserved VP2 residue K140 of KREMEN1-dependent enteroviruses is critical for virus-receptor interactions and viral infection.mBio. 2025 Feb 5;16(2):e0304024. doi: 10.1128/mbio.03040-24. Epub 2025 Jan 16. mBio. 2025. PMID: 39817751 Free PMC article.
-
Pathological characteristics of a murine oral coxsackievirus A10 infection model.J Virol. 2025 Jul 22;99(7):e0093725. doi: 10.1128/jvi.00937-25. Epub 2025 Jul 1. J Virol. 2025. PMID: 40590495 Free PMC article.
References
-
- Ji T, Guo Y, Huang W, Shi Y, Xu Y, Tong W, et al.. The emerging sub-genotype C2 of CoxsackievirusA10 Associated with Hand, Foot and Mouth Disease extensively circulating in mainland of China. Sci Rep. 2018;8(1):13357. Epub 2018/09/08. doi: 10.1038/s41598-018-31616-x [pii]. ; PubMed Central PMCID: PMC6127217. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
