

Secondary structures that regulate mRNA translation provide insights for ASO-mediated modulation of cardiac hypertrophy
- PMID: 37789015
- PMCID: PMC10547706
- DOI: 10.1038/s41467-023-41799-1
Secondary structures that regulate mRNA translation provide insights for ASO-mediated modulation of cardiac hypertrophy
Abstract
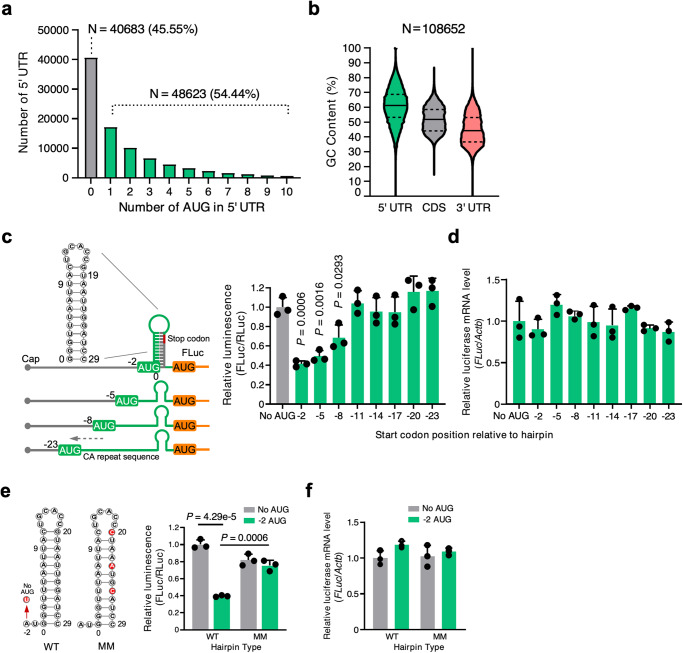
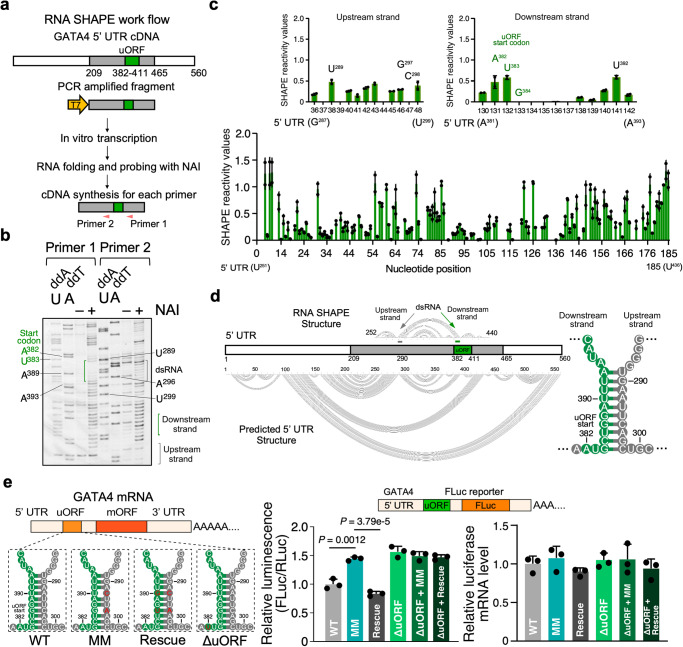
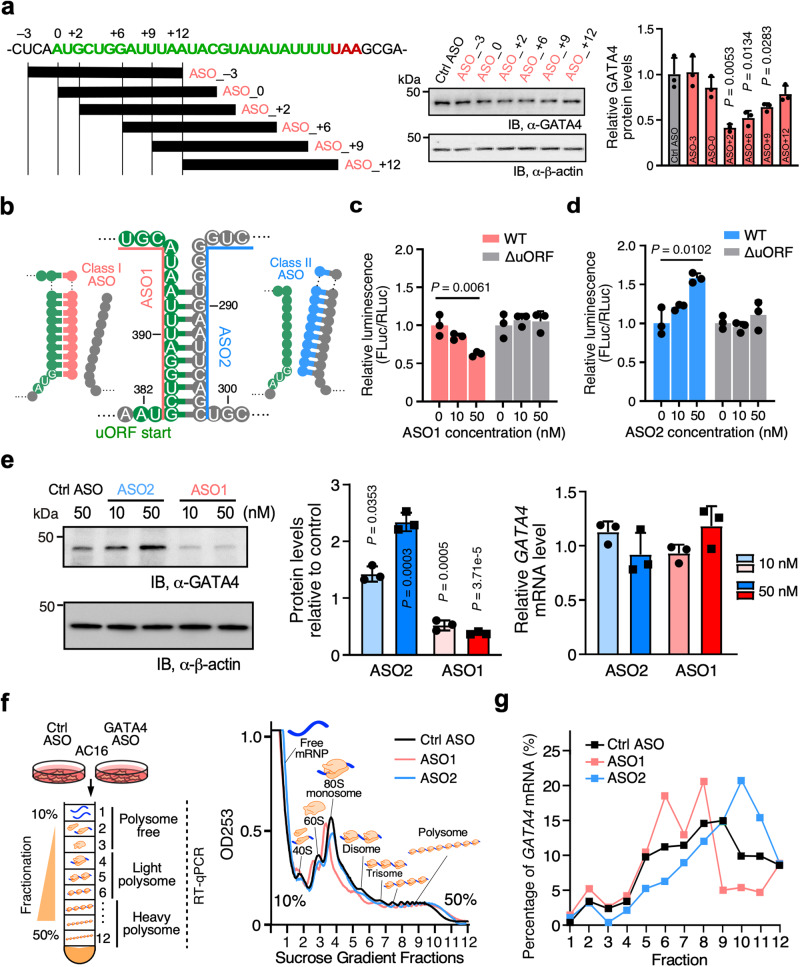
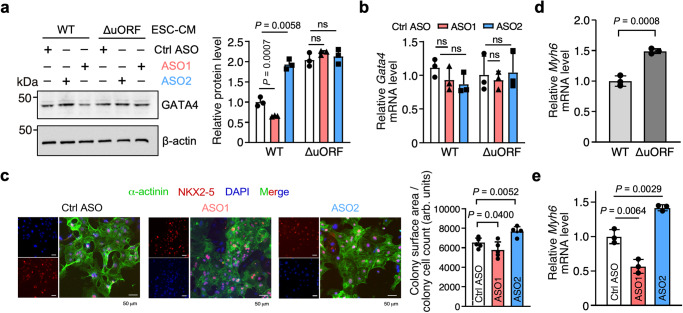
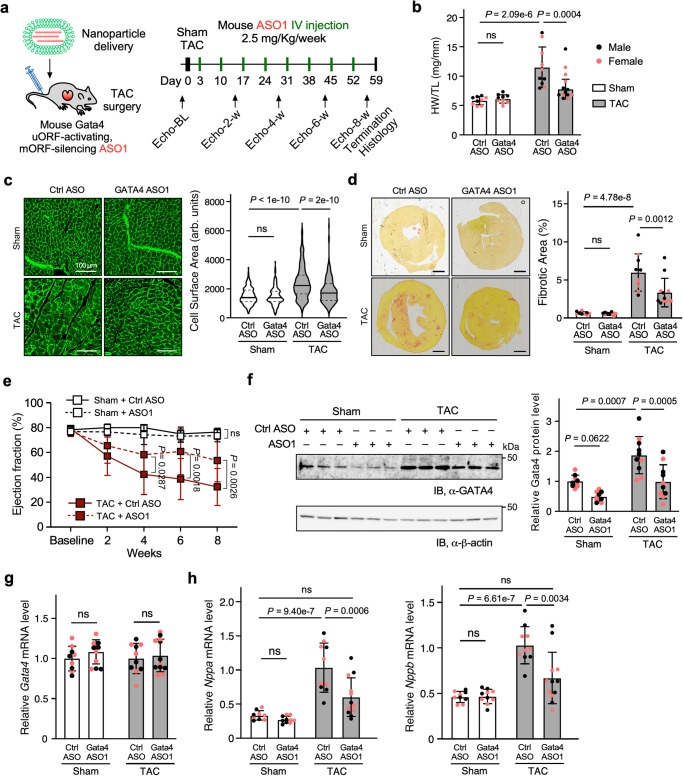
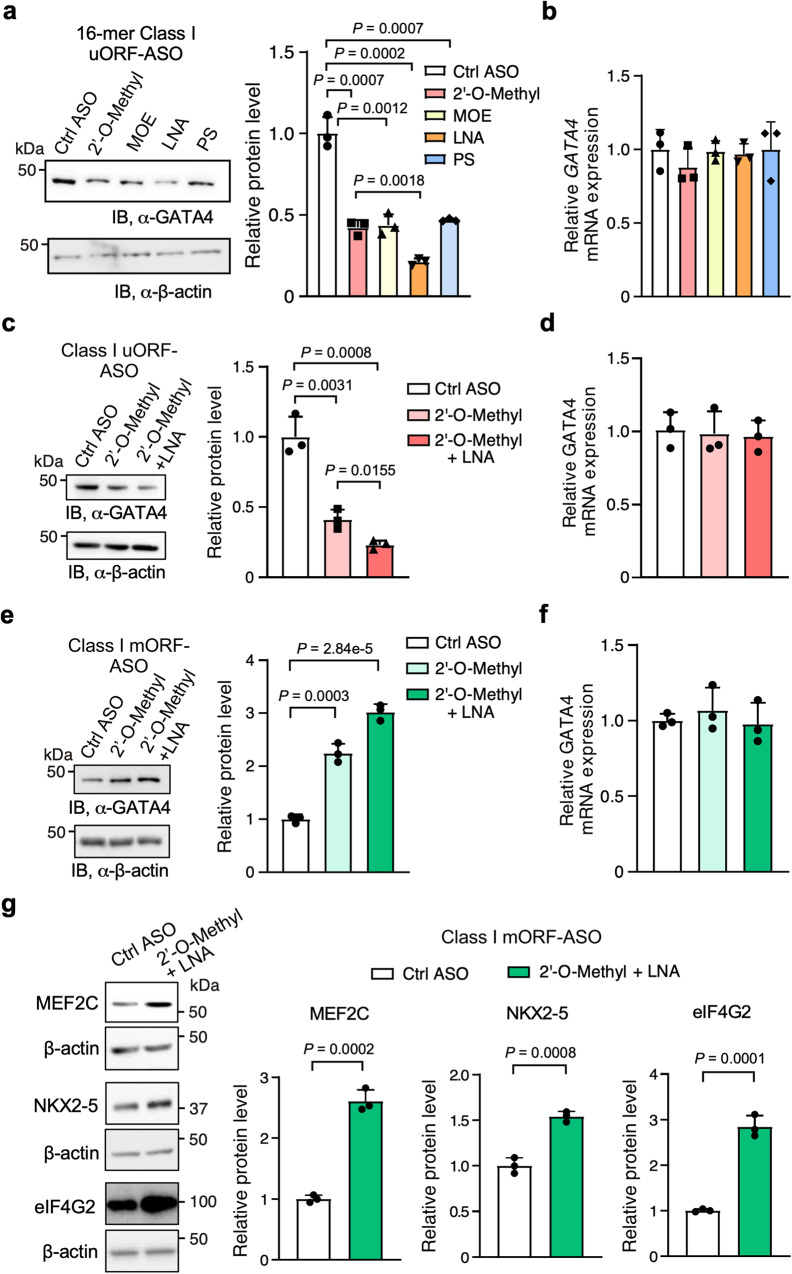
Translation of upstream open reading frames (uORFs) typically abrogates translation of main (m)ORFs. The molecular mechanism of uORF regulation in cells is not well understood. Here, we data-mined human and mouse heart ribosome profiling analyses and identified a double-stranded RNA (dsRNA) structure within the GATA4 uORF that cooperates with the start codon to augment uORF translation and inhibits mORF translation. A trans-acting RNA helicase DDX3X inhibits the GATA4 uORF-dsRNA activity and modulates the translational balance of uORF and mORF. Antisense oligonucleotides (ASOs) that disrupt this dsRNA structure promote mORF translation, while ASOs that base-pair immediately downstream (i.e., forming a bimolecular double-stranded region) of either the uORF or mORF start codon enhance uORF or mORF translation, respectively. Human cardiomyocytes and mice treated with a uORF-enhancing ASO showed reduced cardiac GATA4 protein levels and increased resistance to cardiomyocyte hypertrophy. We further show the broad utility of uORF-dsRNA- or mORF-targeting ASO to regulate mORF translation for other mRNAs. This work demonstrates that the uORF-dsRNA element regulates the translation of multiple mRNAs as a generalizable translational control mechanism. Moreover, we develop a valuable strategy to alter protein expression and cellular phenotypes by targeting or generating dsRNA downstream of a uORF or mORF start codon.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Secondary structures that regulate mRNA translation provide insights for ASO-mediated modulation of cardiac hypertrophy.bioRxiv [Preprint]. 2023 Jun 15:2023.06.15.545153. doi: 10.1101/2023.06.15.545153. bioRxiv. 2023. Update in: Nat Commun. 2023 Oct 3;14(1):6166. doi: 10.1038/s41467-023-41799-1. PMID: 37397986 Free PMC article. Updated. Preprint.
Similar articles
-
Secondary structures that regulate mRNA translation provide insights for ASO-mediated modulation of cardiac hypertrophy.bioRxiv [Preprint]. 2023 Jun 15:2023.06.15.545153. doi: 10.1101/2023.06.15.545153. bioRxiv. 2023. Update in: Nat Commun. 2023 Oct 3;14(1):6166. doi: 10.1038/s41467-023-41799-1. PMID: 37397986 Free PMC article. Updated. Preprint.
-
Translational regulation by uORFs and start codon selection stringency.Genes Dev. 2023 Jun 1;37(11-12):474-489. doi: 10.1101/gad.350752.123. Epub 2023 Jul 11. Genes Dev. 2023. PMID: 37433636 Free PMC article. Review.
-
Temperature-dependent regulation of upstream open reading frame translation in S. cerevisiae.BMC Biol. 2019 Dec 6;17(1):101. doi: 10.1186/s12915-019-0718-5. BMC Biol. 2019. PMID: 31810458 Free PMC article.
-
Specific Increase of Protein Levels by Enhancing Translation Using Antisense Oligonucleotides Targeting Upstream Open Frames.Adv Exp Med Biol. 2017;983:129-146. doi: 10.1007/978-981-10-4310-9_9. Adv Exp Med Biol. 2017. PMID: 28639196 Review.
-
Genome-wide identification of Arabidopsis non-AUG-initiated upstream ORFs with evolutionarily conserved regulatory sequences that control protein expression levels.Plant Mol Biol. 2023 Jan;111(1-2):37-55. doi: 10.1007/s11103-022-01309-1. Epub 2022 Aug 31. Plant Mol Biol. 2023. PMID: 36044152
Cited by
-
Protein binding in an mRNA 5'-UTR sterically hinders translation.RNA. 2025 Jan 22;31(2):143-149. doi: 10.1261/rna.080136.124. RNA. 2025. PMID: 39662963
-
Value of Bioinformatics Models for Predicting Translational Control of Angiogenesis.Circ Res. 2025 May 9;136(10):1147-1165. doi: 10.1161/CIRCRESAHA.125.325438. Epub 2025 May 8. Circ Res. 2025. PMID: 40339045 Review.
-
uORF-targeting steric block antisense oligonucleotides do not reproducibly increase RNASEH1 expression.Mol Ther Nucleic Acids. 2024 Nov 28;36(1):102406. doi: 10.1016/j.omtn.2024.102406. eCollection 2025 Mar 11. Mol Ther Nucleic Acids. 2024. PMID: 39759875 Free PMC article.
-
RNA dysregulation in neurodegenerative diseases.EMBO J. 2025 Feb;44(3):613-638. doi: 10.1038/s44318-024-00352-6. Epub 2025 Jan 9. EMBO J. 2025. PMID: 39789319 Free PMC article. Review.
-
Newborn apical resection preserves the proliferative capacity of cardiomyocytes located throughout the left ventricle.Stem Cells. 2025 May 15;43(5):sxaf018. doi: 10.1093/stmcls/sxaf018. Stem Cells. 2025. PMID: 40229986
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
