

Yang cycle enzyme DEP1: its moonlighting functions in PSI and ROS production during leaf senescence
- PMID: 37789483
- PMCID: PMC10514949
- DOI: 10.1186/s43897-022-00031-2
Yang cycle enzyme DEP1: its moonlighting functions in PSI and ROS production during leaf senescence
Abstract
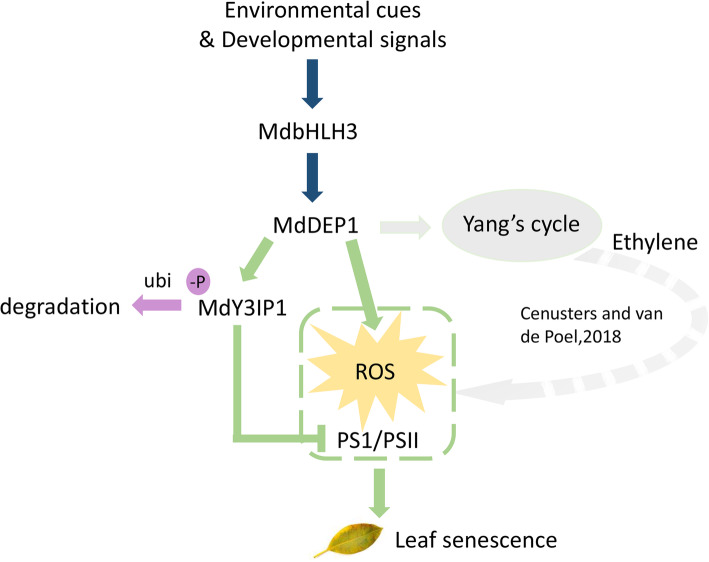
Ethylene-mediated leaf senescence and the compromise of photosynthesis are closely associated but the underlying molecular mechanism is a mystery. Here we reported that apple DEHYDRATASE-ENOLASE-PHOSPHATASE-COMPLEX1 (MdDEP1), initially characterized to its enzymatic function in the recycling of the ethylene precursor SAM, plays a role in the regulation of photosystem I (PSI) activity, activating reactive oxygen species (ROS) homeostasis, and negatively regulating the leaf senescence. A series of Y2H, Pull-down, CO-IP and Cell-free degradation biochemical assays showed that MdDEP1 directly interacts with and dephosphorylates the nucleus-encoded thylakoid protein MdY3IP1, leading to the destabilization of MdY3IP1, reduction of the PSI activity, and the overproduction of ROS in plant cells. These findings elucidate a novel mechanism that the two pathways intersect at MdDEP1 due to its moonlighting role in destabilizing MdY3IP1, and synchronize ethylene-mediated leaf senescence and the compromise of photosynthesis.
Keywords: DEHYDRATASE-ENOLASE-PHOSPHATASE-COMPLEX1; Leaf senescence; Photosynthesis; Photosystem I; ROS; Yang cycle.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures






References
Grants and funding
LinkOut - more resources
Full Text Sources