

This is a preprint.
Computational design of sequence-specific DNA-binding proteins
- PMID: 37790440
- PMCID: PMC10542524
- DOI: 10.1101/2023.09.20.558720
Computational design of sequence-specific DNA-binding proteins
Update in
-
Computational design of sequence-specific DNA-binding proteins.Nat Struct Mol Biol. 2025 Nov;32(11):2252-2261. doi: 10.1038/s41594-025-01669-4. Epub 2025 Sep 12. Nat Struct Mol Biol. 2025. PMID: 40940539 Free PMC article.
Abstract
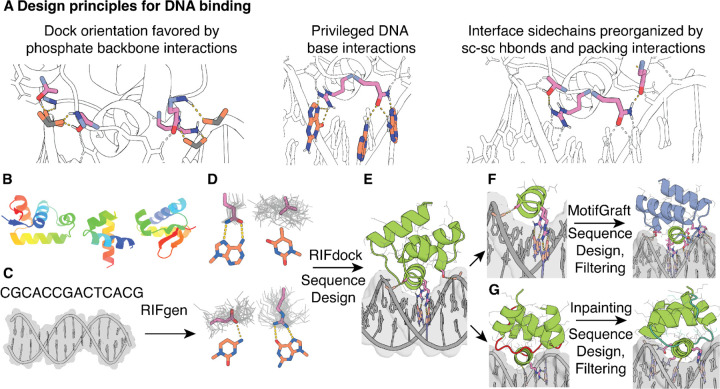
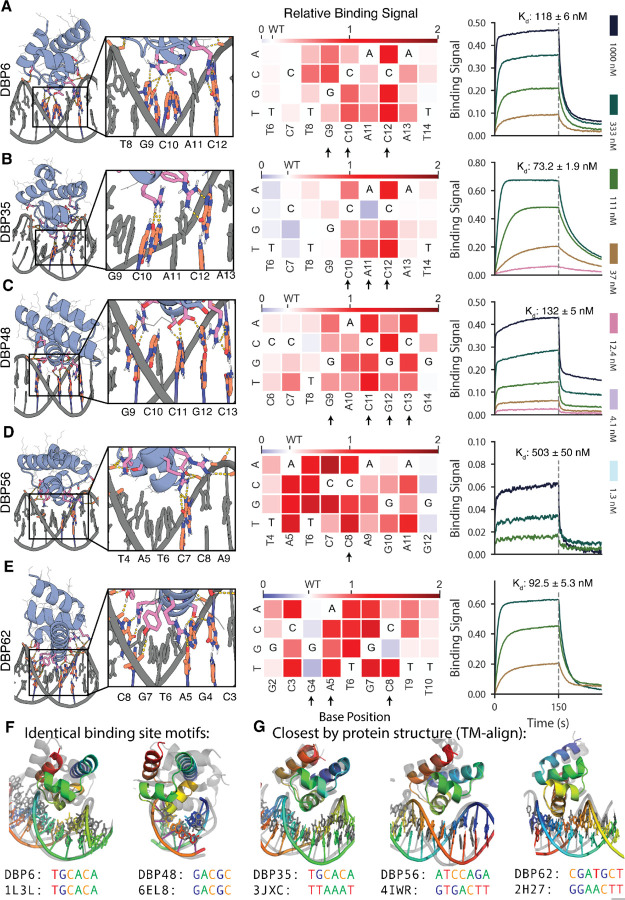
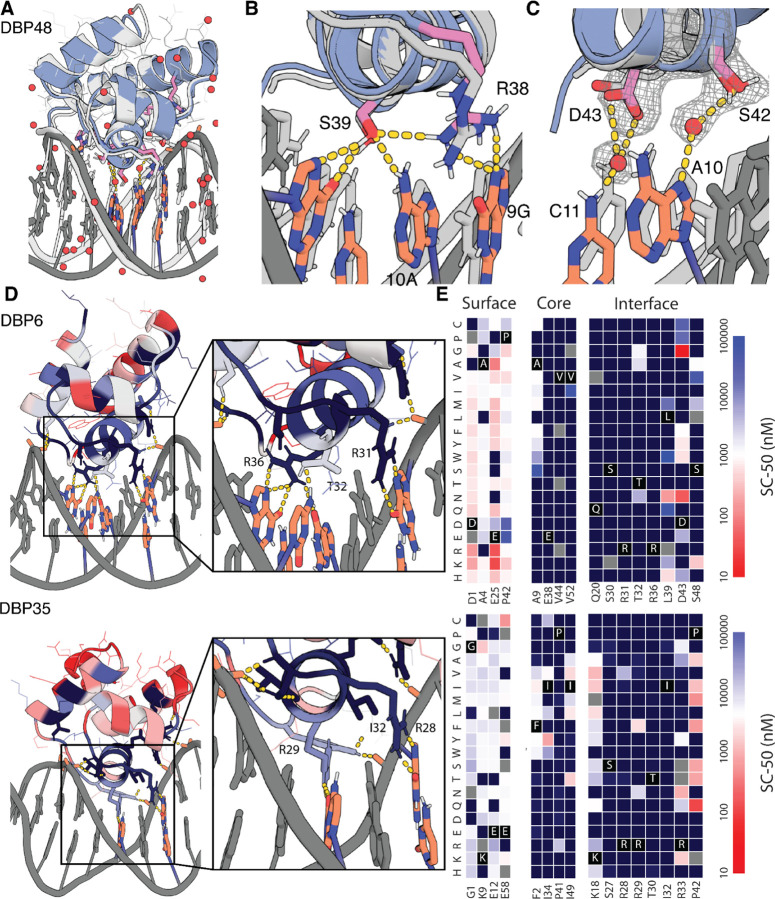
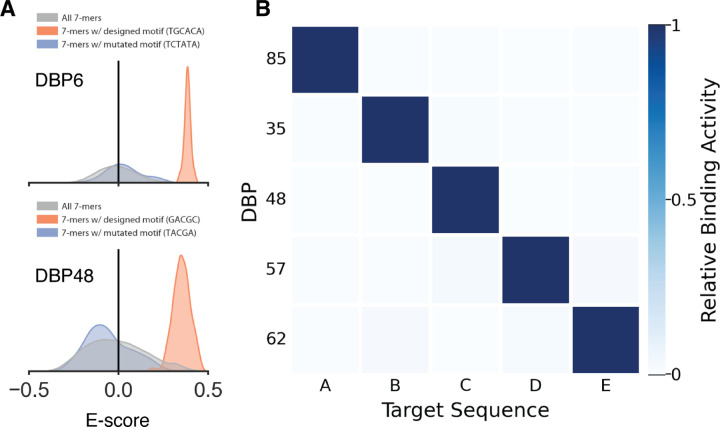
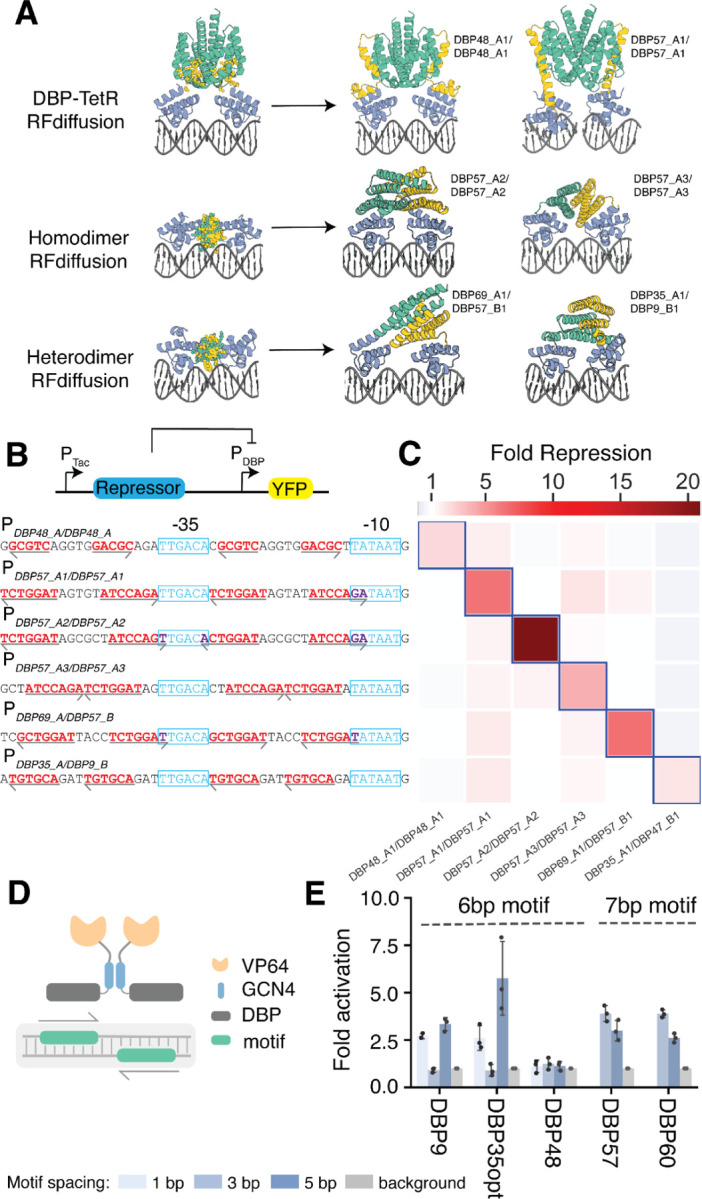
Sequence-specific DNA-binding proteins (DBPs) play critical roles in biology and biotechnology, and there has been considerable interest in the engineering of DBPs with new or altered specificities for genome editing and other applications. While there has been some success in reprogramming naturally occurring DBPs using selection methods, the computational design of new DBPs that recognize arbitrary target sites remains an outstanding challenge. We describe a computational method for the design of small DBPs that recognize specific target sequences through interactions with bases in the major groove, and employ this method in conjunction with experimental screening to generate binders for 5 distinct DNA targets. These binders exhibit specificity closely matching the computational models for the target DNA sequences at as many as 6 base positions and affinities as low as 30-100 nM. The crystal structure of a designed DBP-target site complex is in close agreement with the design model, highlighting the accuracy of the design method. The designed DBPs function in both Escherichia coli and mammalian cells to repress and activate transcription of neighboring genes. Our method is a substantial step towards a general route to small and hence readily deliverable sequence-specific DBPs for gene regulation and editing.
Conflict of interest statement
Competing interests. C.G., R.P., R.M., C.N., F.D., and D.B. are co-inventors on a provisional patent application that incorporates discoveries described in this manuscript.
Figures
References
-
- Wilken M. S., Ciarlo C., Pearl J., Schanzer E., Liao H., Biber B. V., Queitsch K., Bloom J., Federation A., Acosta R., Vong S., Otterman E., Dunn D., Wang H., Zrazhevskiy P., Nandakumar V., Bates D., Sandstrom R., Urnov F. D., Funnell A., Green S., Stamatoyannopoulos J. A., Quantitative dialing of gene expression via precision targeting of KRAB repressor. bioRxiv (2020), doi: 10.1101/2020.02.19.956730. - DOI
-
- Cao L., Coventry B., Goreshnik I., Huang B., Sheffler W., Park J. S., Jude K. M., Marković I., Kadam R. U., Verschueren K. H. G., Verstraete K., Walsh S. T. R., Bennett N., Phal A., Yang A., Kozodoy L., DeWitt M., Picton L., Miller L., Strauch E.-M., DeBouver N. D., Pires A., Bera A. K., Halabiya S., Hammerson B., Yang W., Bernard S., Stewart L., Wilson I. A., Ruohola-Baker H., Schlessinger J., Lee S., Savvides S. N., Garcia K. C., Baker D., Design of protein-binding proteins from the target structure alone. Nature. 605, 551–560 (2022). - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials