

This is a preprint.
Loss of Bmp2 impairs odontogenesis via dysregulating pAkt/pErk/GCN5/Dlx3/Sp7
- PMID: 37790473
- PMCID: PMC10543288
- DOI: 10.21203/rs.3.rs-3299295/v1
Loss of Bmp2 impairs odontogenesis via dysregulating pAkt/pErk/GCN5/Dlx3/Sp7
Abstract
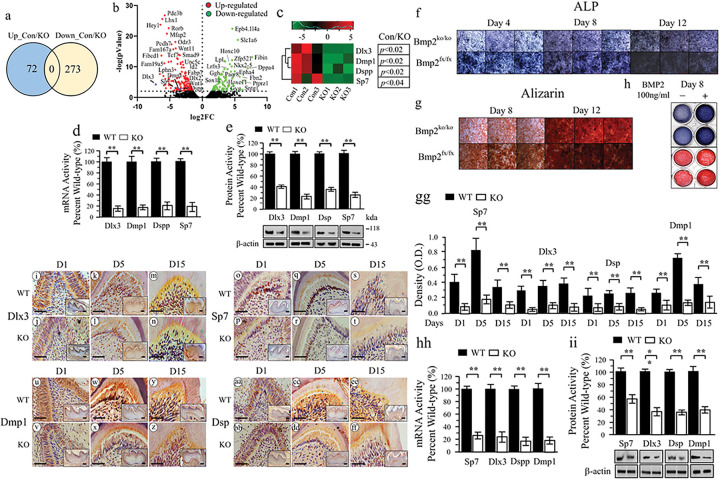
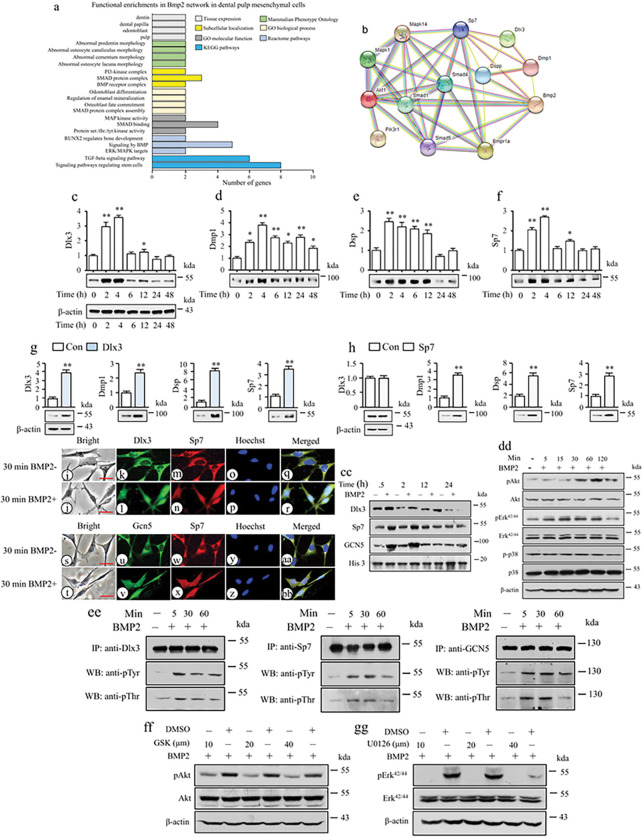
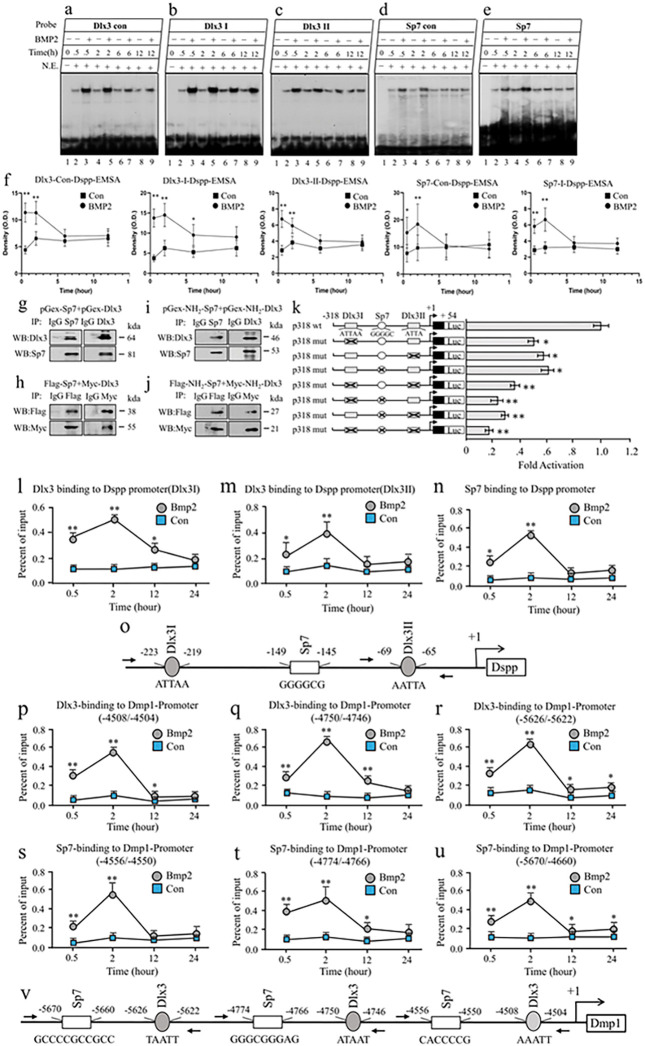
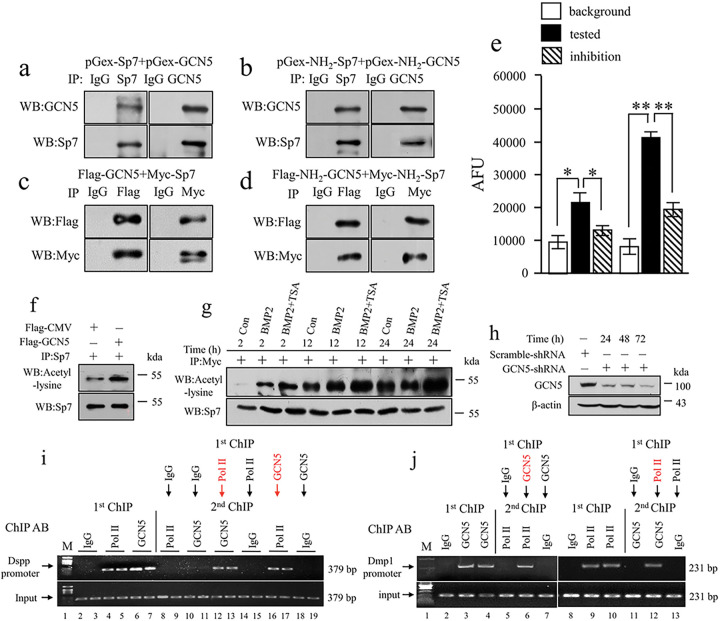
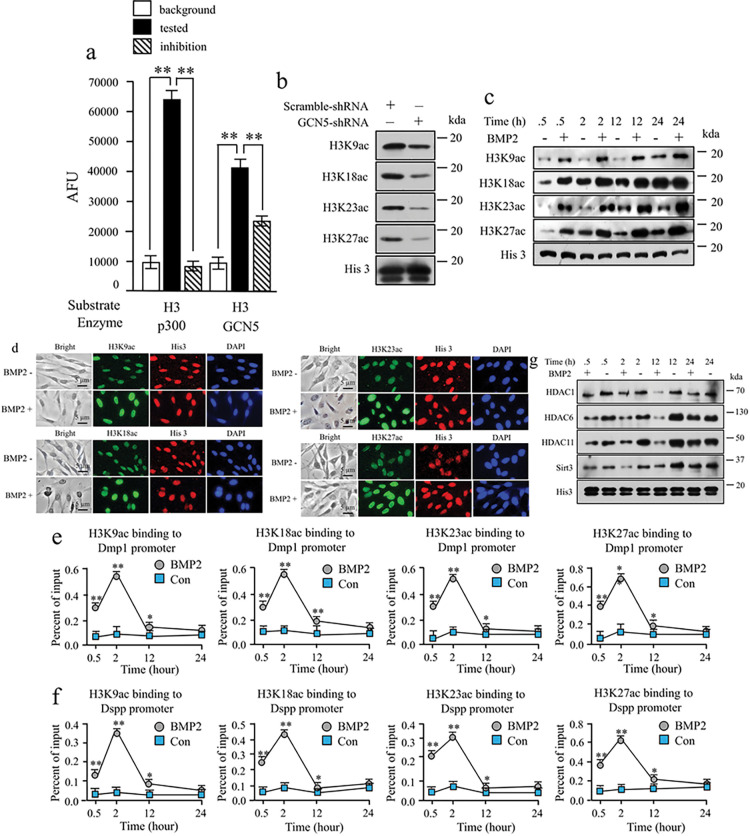
BMP2 signaling plays a pivotal role in odontoblast differentiation and maturation during odontogenesis. Teeth lacking Bmp2 exhibit a morphology reminiscent of dentinogenesis imperfecta (DGI), associated with mutations in dentin matrix protein 1 (DMP1) and dentin sialophosphoprotein (DSPP) genes. Mechanisms by which BMP2 signaling influences expressions of DSPP and DMP1 and contributes to DGI remain elusive. To study the roles of BMP2 in dentin development, we generated Bmp2 conditional knockout (cKO) mice. Through a comprehensive approach involving RNA-seq, immunohistochemistry, promoter activity, ChIP, and Re-ChIP, we investigated downstream targets of Bmp2. Notably, the absence of Bmp2 in cKO mice led to dentin insufficiency akin to DGI. Disrupted Bmp2 signaling was linked to decreased expression of Dspp and Dmp1, as well as alterations in intracellular translocation of transcription factors Dlx3 and Sp7. Intriguingly, upregulation of Dlx3, Dmp1, Dspp, and Sp7, driven by BMP2, fostered differentiation of dental mesenchymal cells and biomineralization. Mechanistically, BMP2 induced phosphorylation of Dlx3, Sp7, and histone acetyltransferase GCN5 at Thr and Tyr residues, mediated by Akt and Erk42/44 kinases. This phosphorylation facilitated protein nuclear translocation, promoting interactions between Sp7 and Dlx3, as well as with GCN5 on Dspp and Dmp1 promoters. The synergy between Dlx3 and Sp7 bolstered transcription of Dspp and Dmp1. Notably, BMP2-driven GCN5 acetylated Sp7 and histone H3, while also recruiting RNA polymerase II to Dmp1 and Dspp chromatins, enhancing their transcriptions. Intriguingly, BMP2 suppressed the expression of histone deacetylases. we unveil hitherto uncharted involvement of BMP2 in dental cell differentiation and dentine development through pAkt/pErk42/44/Dlx3/Sp7/GCN5/Dspp/Dmp1.
Keywords: Bone morphogenetic protein 2; Chromatin remodeling; Dentine matrix protein 1 (DMP1); Dentine sialophosphoprotein (DSPP); Dentinogenesis; Dentinogenesis imperfecta; Transcriptional factors; histone acetyltransferases.
Conflict of interest statement
Conflict of interests: The authors declare no competing interests.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
