

This is a preprint.
Rare variation in noncoding regions with evolutionary signatures contributes to autism spectrum disorder risk
- PMID: 37790480
- PMCID: PMC10543033
- DOI: 10.1101/2023.09.19.23295780
Rare variation in noncoding regions with evolutionary signatures contributes to autism spectrum disorder risk
Update in
-
Rare variation in non-coding regions with evolutionary signatures contributes to autism spectrum disorder risk.Cell Genom. 2024 Aug 14;4(8):100609. doi: 10.1016/j.xgen.2024.100609. Epub 2024 Jul 16. Cell Genom. 2024. PMID: 39019033 Free PMC article.
Abstract
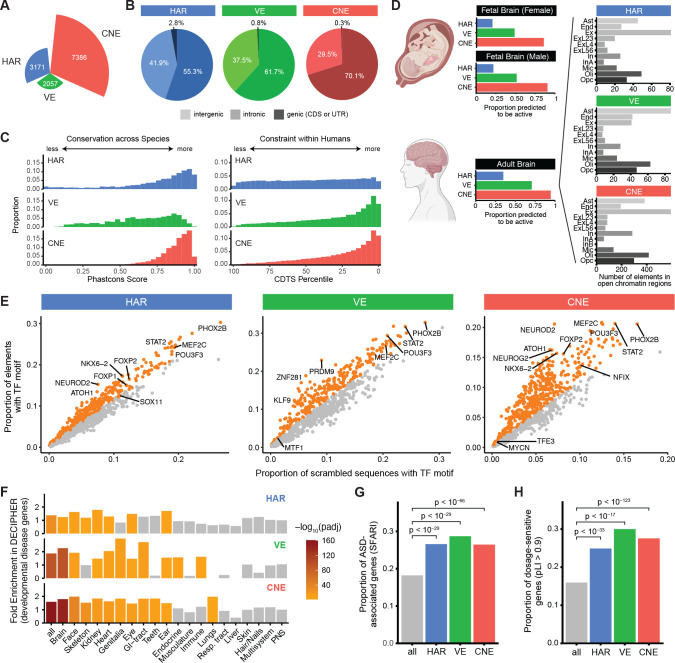
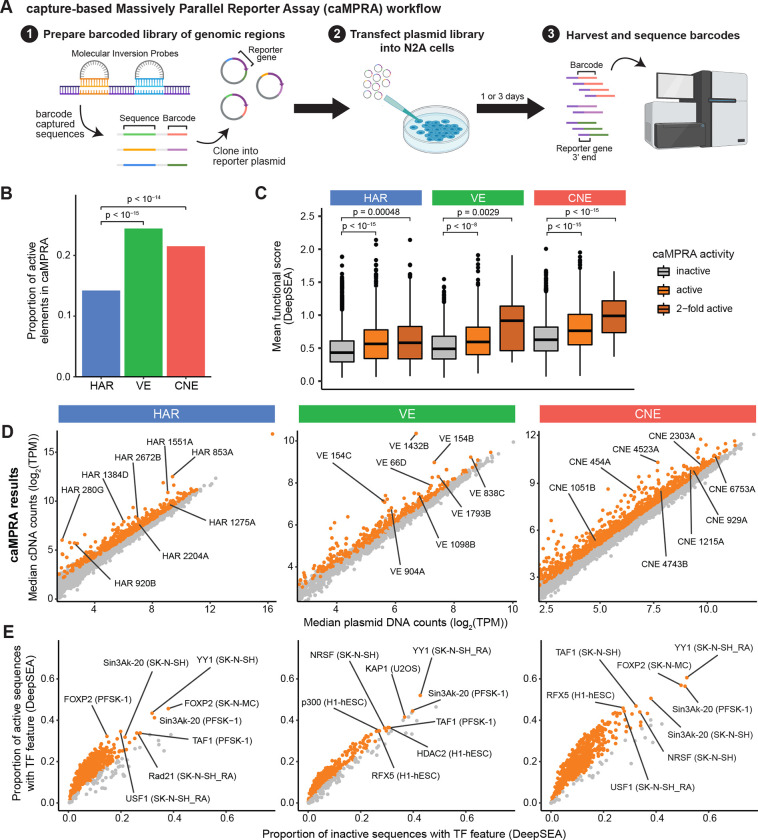
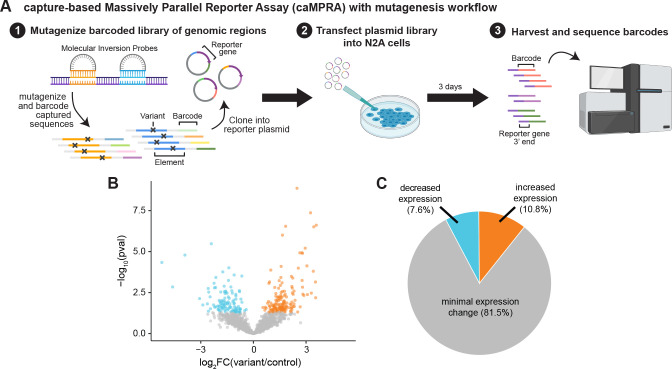
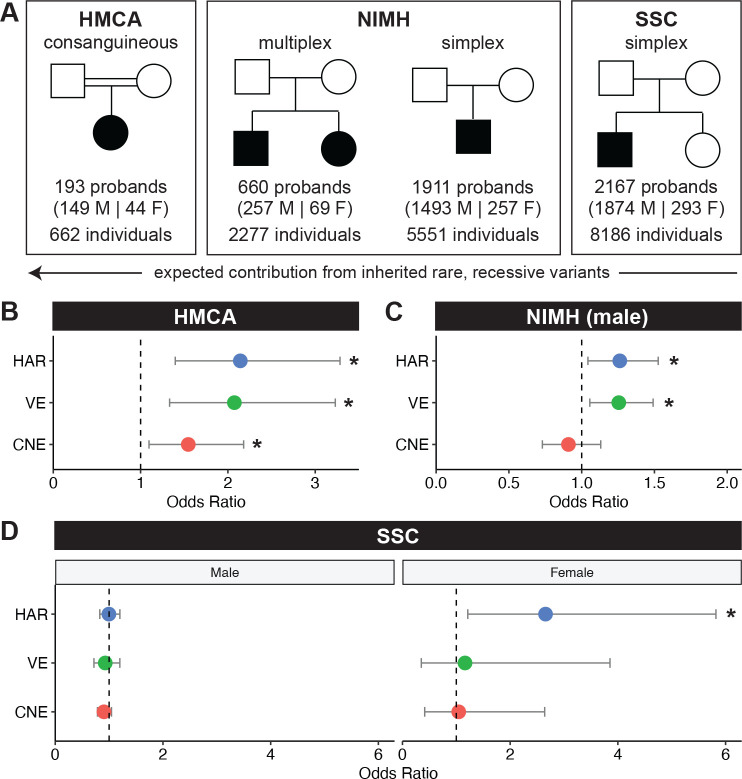
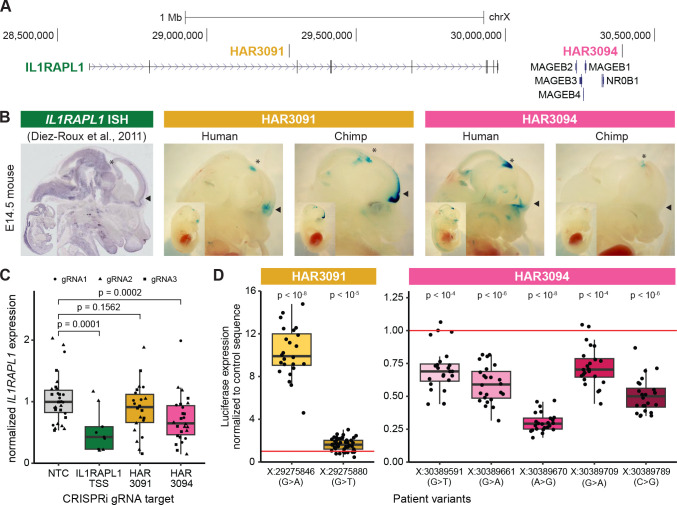
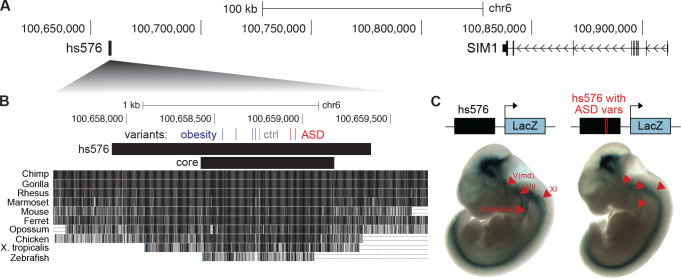
Little is known about the role of noncoding regions in the etiology of autism spectrum disorder (ASD). We examined three classes of noncoding regions: Human Accelerated Regions (HARs), which show signatures of positive selection in humans; experimentally validated neural Vista Enhancers (VEs); and conserved regions predicted to act as neural enhancers (CNEs). Targeted and whole genome analysis of >16,600 samples and >4900 ASD probands revealed that likely recessive, rare, inherited variants in HARs, VEs, and CNEs substantially contribute to ASD risk in probands whose parents share ancestry, which enriches for recessive contributions, but modestly, if at all, in simplex family structures. We identified multiple patient variants in HARs near IL1RAPL1 and in a VE near SIM1 and showed that they change enhancer activity. Our results implicate both human-evolved and evolutionarily conserved noncoding regions in ASD risk and suggest potential mechanisms of how changes in regulatory regions can modulate social behavior.
Keywords: Human Accelerated Regions; IL1RAPL1; SIM1; Vista Enhancers; autism spectrum disorder; caMPRA; consanguineous families; conserved neural enhancers; noncoding regions; recessive variants.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Akula S. K., Marciano J. H., Lim Y., Exposito-Alonso D., Hylton N. K., Hwang G. H., Neil J. E., Dominado N., Bunton-Stasyshyn R. K., Song J. H. T., et al. , 2023. TMEM161B regulates cerebral cortical gyration, Sonic Hedgehog signaling, and ciliary structure in the developing central nervous system. Proceedings of the National Academy of Sciences, 120(4):e2209964120. - PMC - PubMed
-
- Altman D. G., 1991. Practical Statistics for Medical Research. Chapman and Hall, London.