

Ubiquitination of the protocadherin-γA3 variable cytoplasmic domain modulates cell-cell interaction
- PMID: 37791076
- PMCID: PMC10544333
- DOI: 10.3389/fcell.2023.1261048
Ubiquitination of the protocadherin-γA3 variable cytoplasmic domain modulates cell-cell interaction
Abstract
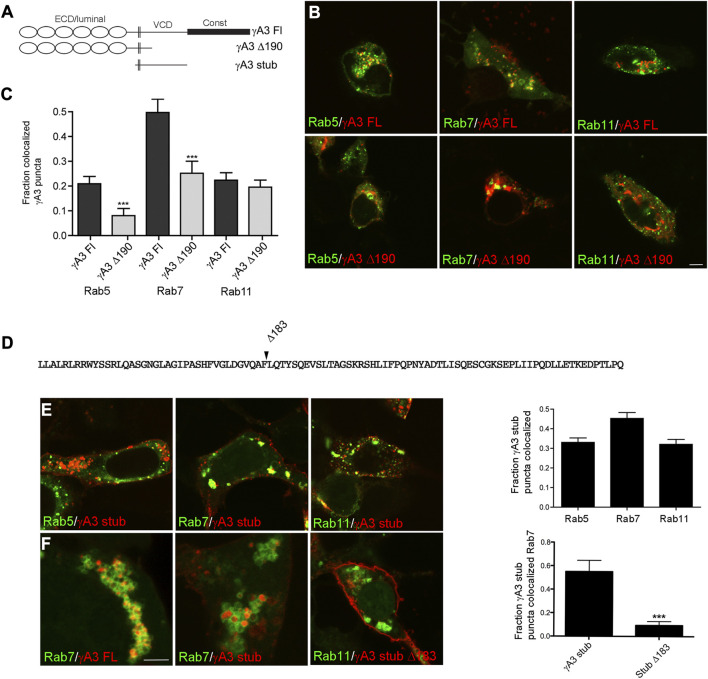
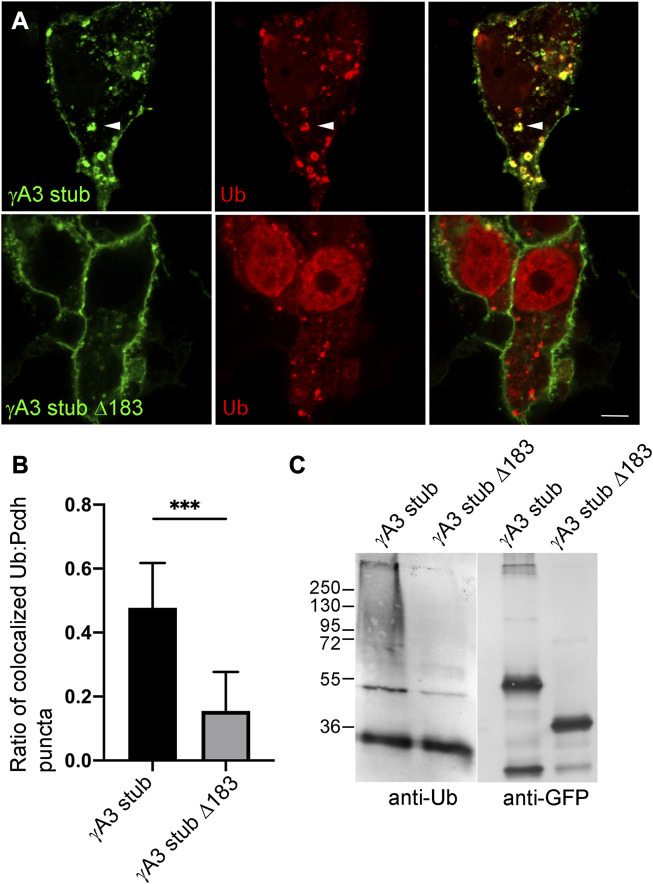
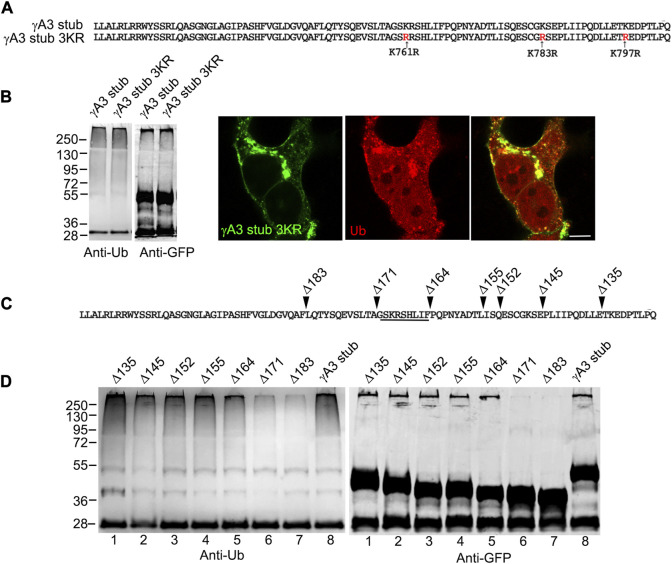
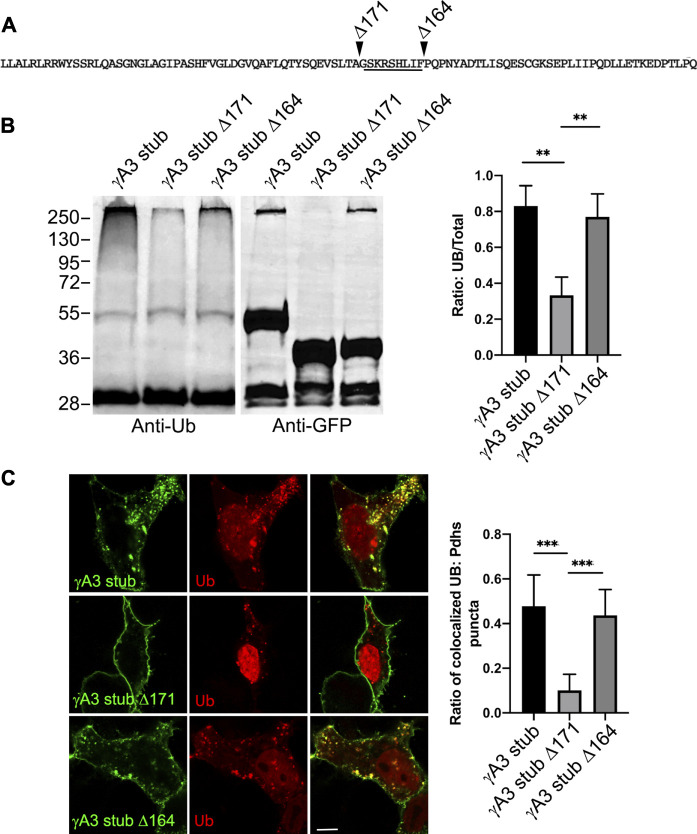
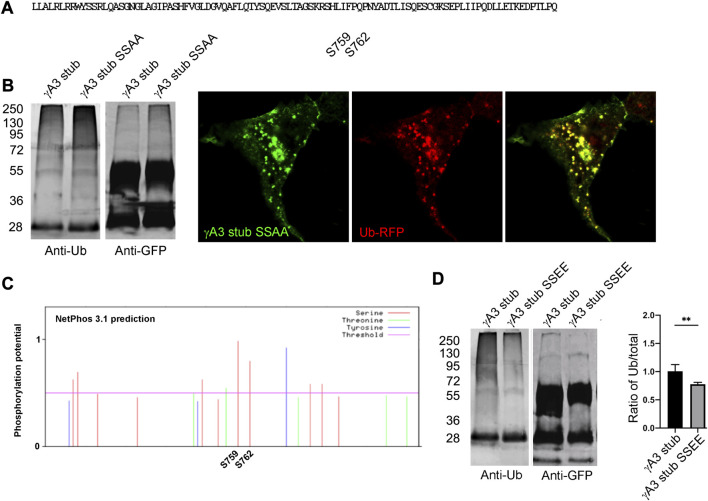
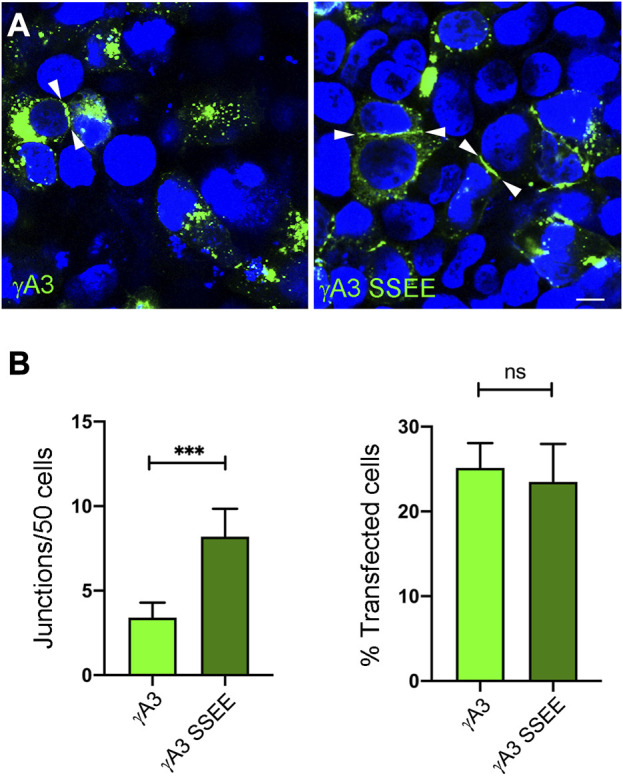
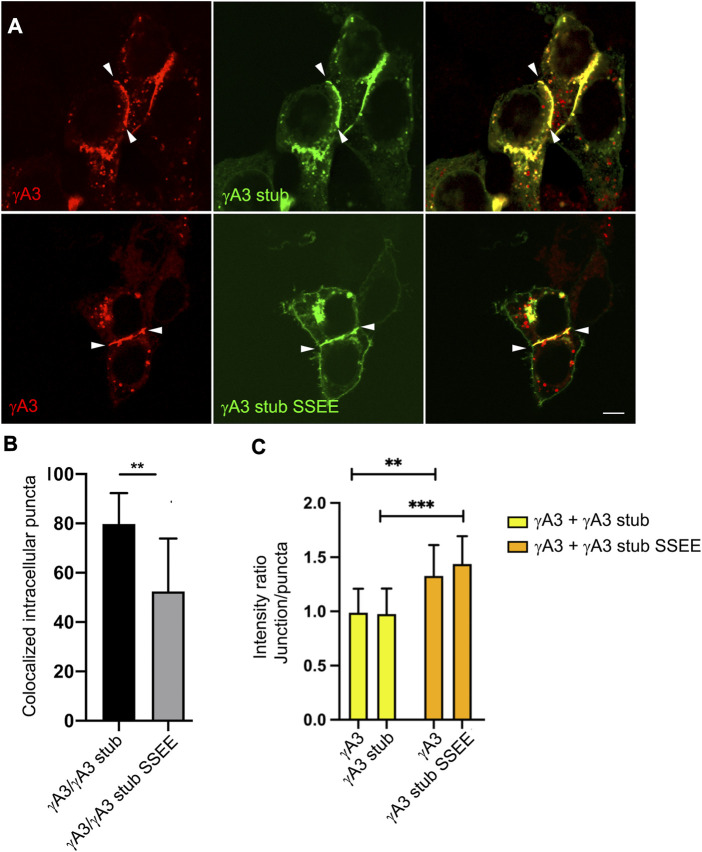
The family of ∼60 clustered protocadherins (Pcdhs) are cell adhesion molecules encoded by a genomic locus that regulates expression of distinct combinations of isoforms in individual neurons resulting in what is thought to be a neural surface "barcode" which mediates same-cell interactions of dendrites, as well as interactions with other cells in the environment. Pcdh mediated same-cell dendrite interactions were shown to result in avoidance while interactions between different cells through Pcdhs, such as between neurons and astrocytes, appear to be stable. The cell biological mechanism of the consequences of Pcdh based adhesion is not well understood although various signaling pathways have been recently uncovered. A still unidentified cytoplasmic regulatory mechanism might contribute to a "switch" between avoidance and adhesion. We have proposed that endocytosis and intracellular trafficking could be part of such a switch. Here we use "stub" constructs consisting of the proximal cytoplasmic domain (lacking the constant carboxy-terminal domain spliced to all Pcdh-γs) of one Pcdh, Pcdh-γA3, to study trafficking. We found that the stub construct traffics primarily to Rab7 positive endosomes very similarly to the full length molecule and deletion of a substantial portion of the carboxy-terminus of the stub eliminates this trafficking. The intact stub was found to be ubiquitinated while the deletion was not and this ubiquitination was found to be at non-lysine sites. Further deletion mapping of the residues required for ubiquitination identified potential serine phosphorylation sites, conserved among Pcdh-γAs, that can reduce ubiquitination when pseudophosphorylated and increase surface expression. These results suggest Pcdh-γA ubiquitination can influence surface expression which may modulate adhesive activity during neural development.
Keywords: cell adhesion; endocytosis; endosome; pseudophosphorylation; self-avoidance.
Copyright © 2023 Ptashnik, LaMassa, Mambetalieva, Schnall, Bucaro and Phillips.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The authors declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision
Figures
References
LinkOut - more resources
Full Text Sources