

Stroma-Mediated Breast Cancer Cell Proliferation Indirectly Drives Chemoresistance by Accelerating Tumor Recovery between Chemotherapy Cycles
- PMID: 37791818
- PMCID: PMC10646478
- DOI: 10.1158/0008-5472.CAN-23-0398
Stroma-Mediated Breast Cancer Cell Proliferation Indirectly Drives Chemoresistance by Accelerating Tumor Recovery between Chemotherapy Cycles
Abstract
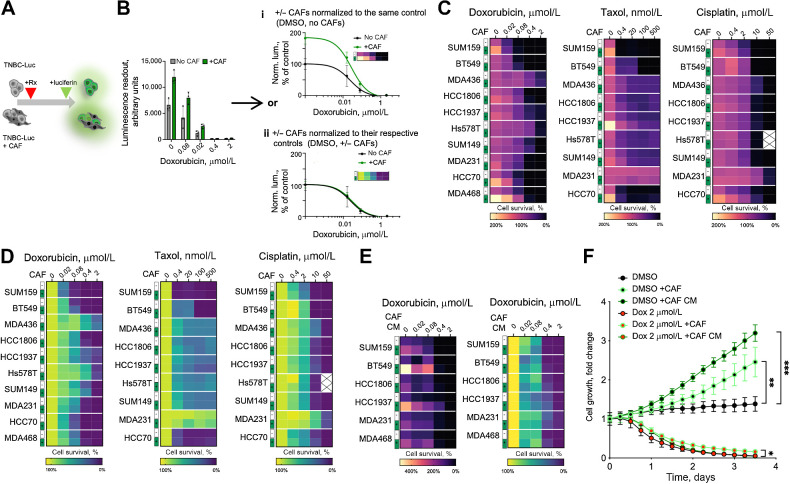
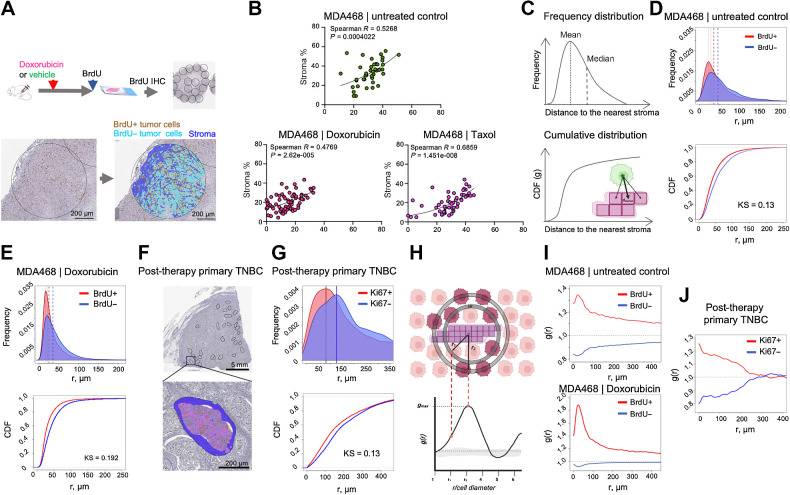
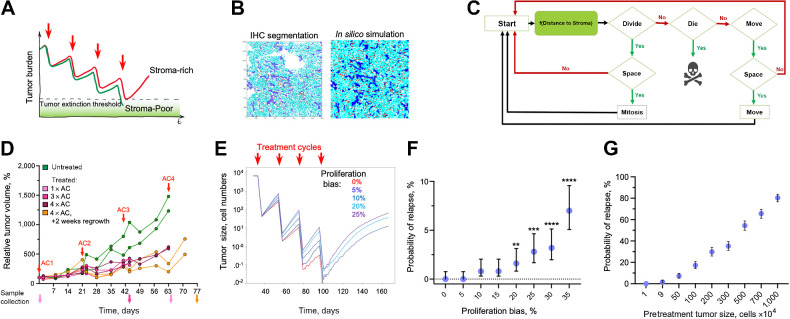
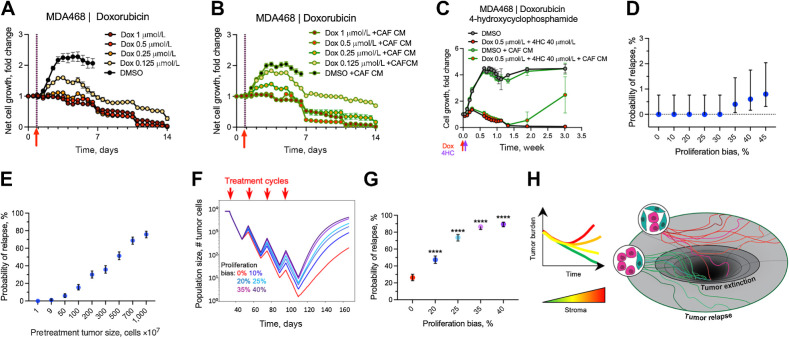
The ability of tumors to survive therapy reflects both cell-intrinsic and microenvironmental mechanisms. Across many cancers, including triple-negative breast cancer (TNBC), a high stroma/tumor ratio correlates with poor survival. In many contexts, this correlation can be explained by the direct reduction of therapy sensitivity induced by stroma-produced paracrine factors. We sought to explore whether this direct effect contributes to the link between stroma and poor responses to chemotherapies. In vitro studies with panels of TNBC cell line models and stromal isolates failed to detect a direct modulation of chemoresistance. At the same time, consistent with prior studies, fibroblast-produced secreted factors stimulated treatment-independent enhancement of tumor cell proliferation. Spatial analyses indicated that proximity to stroma is often associated with enhanced tumor cell proliferation in vivo. These observations suggested an indirect link between stroma and chemoresistance, where stroma-augmented proliferation potentiates the recovery of residual tumors between chemotherapy cycles. To evaluate this hypothesis, a spatial agent-based model of stroma impact on proliferation/death dynamics was developed that was quantitatively parameterized using inferences from histologic analyses and experimental studies. The model demonstrated that the observed enhancement of tumor cell proliferation within stroma-proximal niches could enable tumors to avoid elimination over multiple chemotherapy cycles. Therefore, this study supports the existence of an indirect mechanism of environment-mediated chemoresistance that might contribute to the negative correlation between stromal content and poor therapy outcomes.
Significance: Integration of experimental research with mathematical modeling reveals an indirect microenvironmental chemoresistance mechanism by which stromal cells stimulate breast cancer cell proliferation and highlights the importance of consideration of proliferation/death dynamics. See related commentary by Wall and Echeverria, p. 3667.
©2023 The Authors; Published by the American Association for Cancer Research.
Figures
Comment in
-
Avoiding Extinction: Cancer-Associated Fibroblasts Help Triple-Negative Breast Cancer Outrun Chemotherapy.Cancer Res. 2023 Nov 15;83(22):3667-3669. doi: 10.1158/0008-5472.CAN-23-2770. Cancer Res. 2023. PMID: 37964615
References
-
- Jamieson NB, Carter CR, McKay CJ, Oien KA. Tissue biomarkers for prognosis in pancreatic ductal adenocarcinoma: a systematic review and meta-analysis. Clin Cancer Res 2011;17:3316–31. - PubMed
-
- Graizel D, Zlotogorski-Hurvitz A, Tsesis I, Rosen E, Kedem R, Vered M. Oral cancer-associated fibroblasts predict poor survival: systematic review and meta-analysis. Oral Dis 2020;26:733–44. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
