

Systematic sequence engineering enhances the induction strength of the glucose-regulated GTH1 promoter of Komagataella phaffii
- PMID: 37791854
- PMCID: PMC10639056
- DOI: 10.1093/nar/gkad752
Systematic sequence engineering enhances the induction strength of the glucose-regulated GTH1 promoter of Komagataella phaffii
Abstract
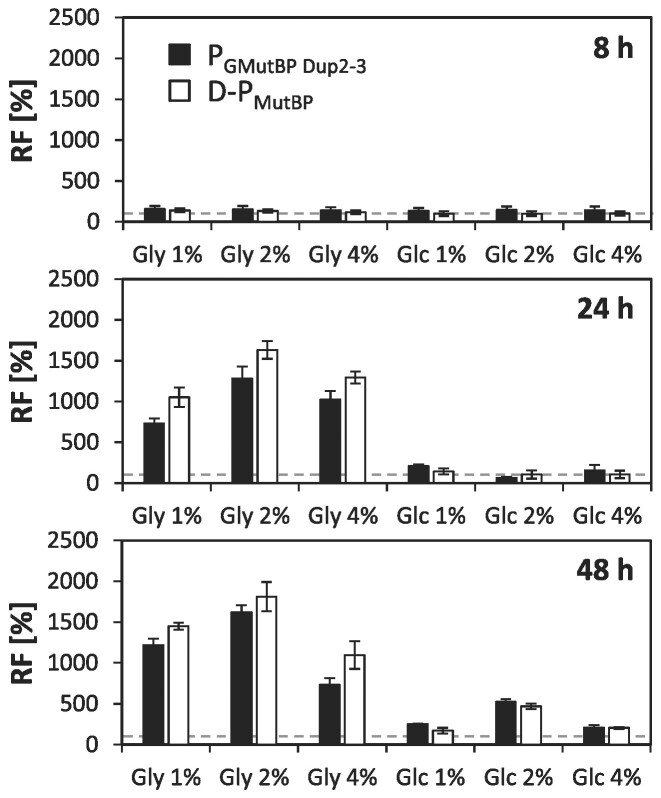
The promoter of the high-affinity glucose transporter Gth1 (PGTH1) is tightly repressed on glucose and glycerol surplus, and strongly induced in glucose-limitation, thus enabling regulated methanol-free production processes in the yeast production host Komagataella phaffii. To further improve this promoter, an intertwined approach of nucleotide diversification through random and rational engineering was pursued. Random mutagenesis and fluorescence activated cell sorting of PGTH1 yielded five variants with enhanced induction strength. Reverse engineering of individual point mutations found in the improved variants identified two single point mutations with synergistic action. Sequential deletions revealed the key promoter segments for induction and repression properties, respectively. Combination of the single point mutations and the amplification of key promoter segments led to a library of novel promoter variants with up to 3-fold higher activity. Unexpectedly, the effect of gaining or losing a certain transcription factor binding site (TFBS) was highly dependent on its context within the promoter. Finally, the applicability of the novel promoter variants for biotechnological production was proven for the secretion of different recombinant model proteins in fed batch cultivation, where they clearly outperformed their ancestors. In addition to advancing the toolbox for recombinant protein production and metabolic engineering of K. phaffii, we discovered single nucleotide positions and correspondingly affected TFBS that distinguish between glycerol- and glucose-mediated repression of the native promoter.
© The Author(s) 2023. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures









Similar articles
-
Advancing recombinant protein expression in Komagataella phaffii: opportunities and challenges.FEMS Yeast Res. 2025 Jan 30;25:foaf010. doi: 10.1093/femsyr/foaf010. FEMS Yeast Res. 2025. PMID: 40074550 Free PMC article. Review.
-
Knock-out of the major regulator Flo8 in Komagataella phaffii results in unique host strain performance for methanol-free recombinant protein production.N Biotechnol. 2024 Dec 25;84:105-114. doi: 10.1016/j.nbt.2024.10.001. Epub 2024 Oct 9. N Biotechnol. 2024. PMID: 39384085
-
Bi-directionalized promoter systems allow methanol-free production of hard-to-express peroxygenases with Komagataella Phaffii.Microb Cell Fact. 2024 Jun 15;23(1):177. doi: 10.1186/s12934-024-02451-9. Microb Cell Fact. 2024. PMID: 38879507 Free PMC article.
-
Metabolic engineering of Komagataella phaffii for synergetic utilization of glucose and glycerol.Yeast. 2022 Jun;39(6-7):412-421. doi: 10.1002/yea.3793. Epub 2022 Jun 16. Yeast. 2022. PMID: 35650013
-
Expanding the promoter toolbox for metabolic engineering of methylotrophic yeasts.Appl Microbiol Biotechnol. 2022 May;106(9-10):3449-3464. doi: 10.1007/s00253-022-11948-5. Epub 2022 May 11. Appl Microbiol Biotechnol. 2022. PMID: 35538374 Review.
Cited by
-
Komagataella phaffii as a Platform for Heterologous Expression of Enzymes Used for Industry.Microorganisms. 2024 Feb 7;12(2):346. doi: 10.3390/microorganisms12020346. Microorganisms. 2024. PMID: 38399750 Free PMC article. Review.
-
Advancing recombinant protein expression in Komagataella phaffii: opportunities and challenges.FEMS Yeast Res. 2025 Jan 30;25:foaf010. doi: 10.1093/femsyr/foaf010. FEMS Yeast Res. 2025. PMID: 40074550 Free PMC article. Review.
-
Harnessing alkaline-pH regulatable promoters for efficient methanol-free expression of enzymes of industrial interest in Komagataella Phaffii.Microb Cell Fact. 2024 Apr 2;23(1):99. doi: 10.1186/s12934-024-02362-9. Microb Cell Fact. 2024. PMID: 38566096 Free PMC article.
References
-
- Cazier A.P., Blazeck J.. Advances in promoter engineering: novel applications and predefined transcriptional control. Biotechnol. J. 2021; 16:e2100239. - PubMed
-
- Xu N., Wei L., Liu J.. Recent advances in the applications of promoter engineering for the optimization of metabolite biosynthesis. World J. Microbiol. Biotechnol. 2019; 35:33. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
