

Topological damping in an ultrafast giant cell
- PMID: 37792511
- PMCID: PMC10576051
- DOI: 10.1073/pnas.2303940120
Topological damping in an ultrafast giant cell
Abstract
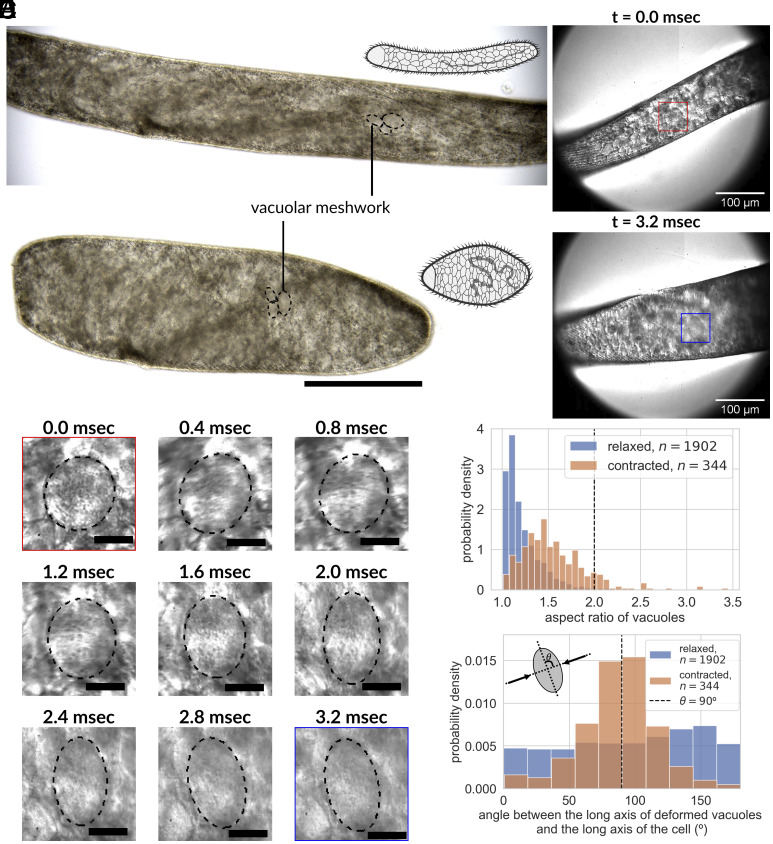
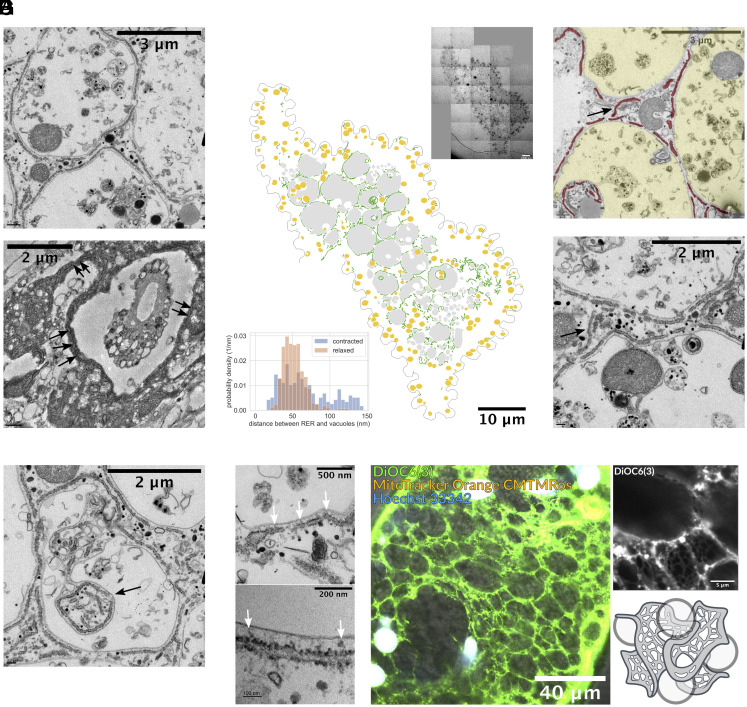
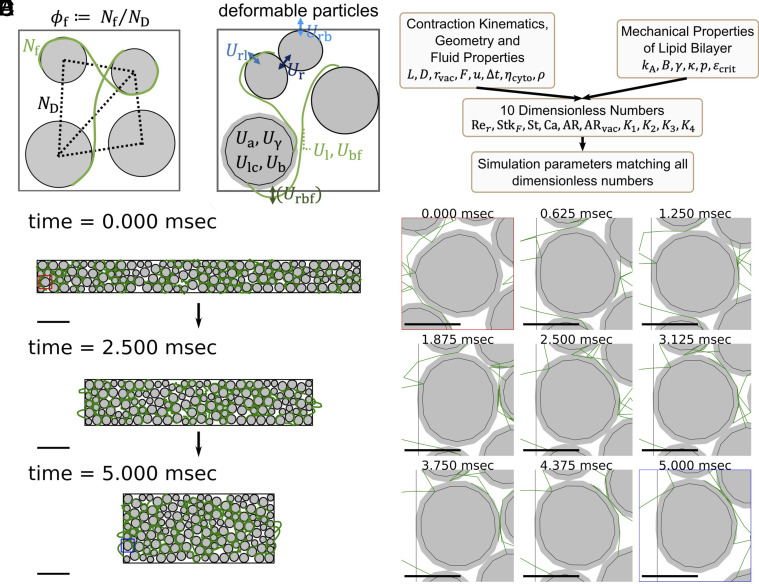
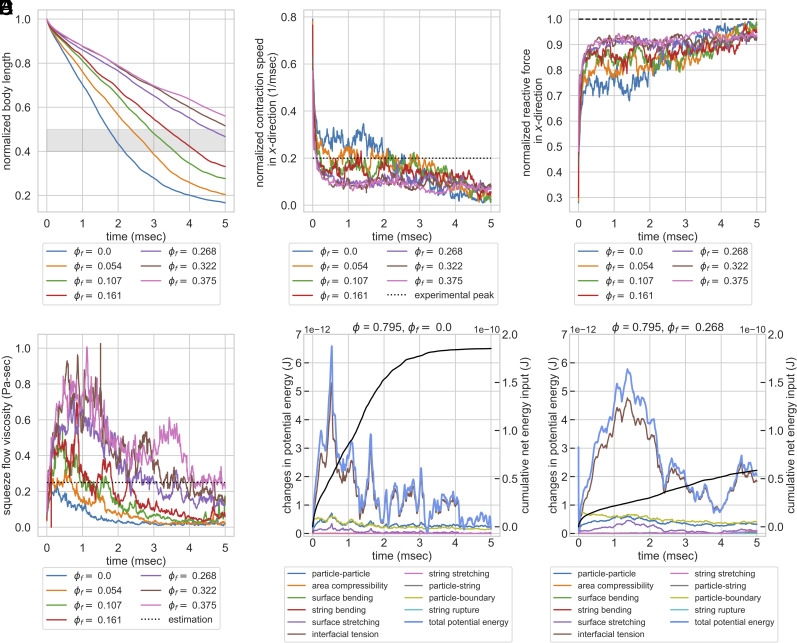
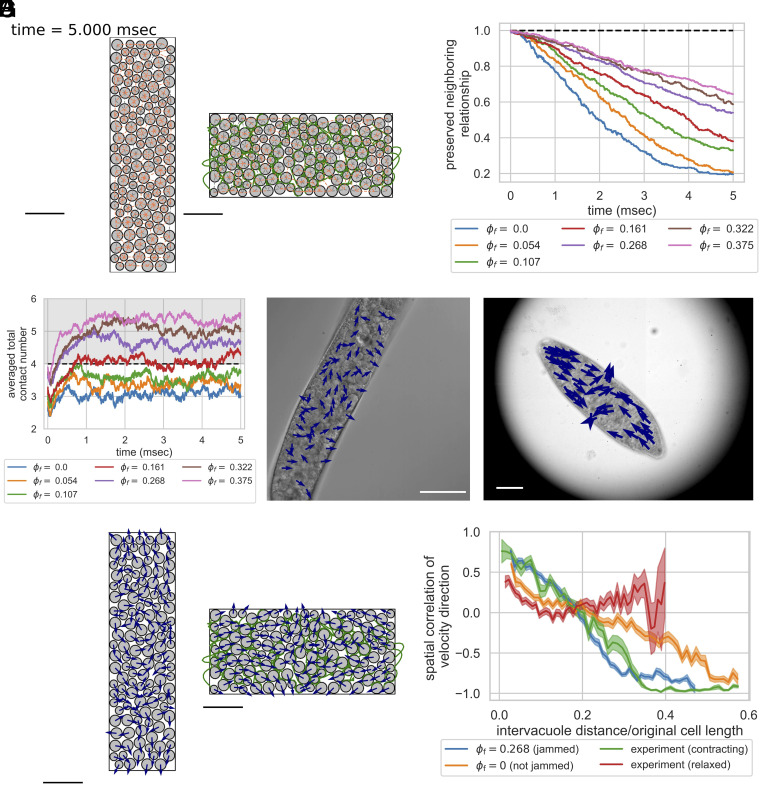
Cellular systems are known to exhibit some of the fastest movements in biology, but little is known as to how single cells can dissipate this energy rapidly and adapt to such large accelerations without disrupting internal architecture. To address this, we investigate Spirostomum ambiguum-a giant cell (1-4 mm in length) well-known to exhibit ultrafast contractions (50% of body length) within 5 ms with a peak acceleration of 15[Formula: see text]. Utilizing transmitted electron microscopy and confocal imaging, we identify an association of rough endoplasmic reticulum (RER) and vacuoles throughout the cell-forming a contiguous fenestrated membrane architecture that topologically entangles these two organelles. A nearly uniform interorganelle spacing of 60 nm is observed between RER and vacuoles, closely packing the entire cell. Inspired by the entangled organelle structure, we study the mechanical properties of entangled deformable particles using a vertex-based model, with all simulation parameters matching 10 dimensionless numbers to ensure dynamic similarity. We demonstrate how entangled deformable particles respond to external loads by an increased viscosity against squeezing and help preserve spatial relationships. Because this enhanced damping arises from the entanglement of two networks incurring a strain-induced jamming transition at subcritical volume fractions, which is demonstrated through the spatial correlation of velocity direction, we term this phenomenon "topological damping." Our findings suggest a mechanical role of RER-vacuolar meshwork as a metamaterial capable of damping an ultrafast contraction event.
Keywords: organelle topology; ultrafast biophysics; vertex model.
Conflict of interest statement
The authors declare no competing interest.
Figures

Similar articles
-
Fishnet mesh of centrin-Sfi1 drives ultrafast calcium-activated contraction of the giant cell Spirostomum ambiguum.bioRxiv [Preprint]. 2024 Nov 8:2024.11.07.622534. doi: 10.1101/2024.11.07.622534. bioRxiv. 2024. PMID: 39574644 Free PMC article. Preprint.
-
Defensive ink pigment processing and secretion in Aplysia californica: concentration and storage of phycoerythrobilin in the ink gland.J Exp Biol. 1998 May;201(Pt 10):1595-613. doi: 10.1242/jeb.201.10.1595. J Exp Biol. 1998. PMID: 9556541
-
Cellular differentiation in moss protonemata: a morphological and experimental study.Ann Bot. 2008 Aug;102(2):227-45. doi: 10.1093/aob/mcn080. Epub 2008 May 27. Ann Bot. 2008. PMID: 18508779 Free PMC article.
-
[Ultrastructural similarities of secretory processes in arterial smooth muscle cells and elastic cartilage chondrocytes. vacuolar apparatus and microtubules (author's transl)].Anat Anz. 1981;150(3):327-34. Anat Anz. 1981. PMID: 7305007 German.
-
Time course of vinblastine-induced autophagocytosis and changes in the endoplasmic reticulum in murine pancreatic acinar cells: a morphometric and biochemical study.Eur J Cell Biol. 1996 Dec;71(4):341-50. Eur J Cell Biol. 1996. PMID: 8980904
Cited by
-
Deformable microlaser force sensing.Light Sci Appl. 2024 Jun 5;13(1):129. doi: 10.1038/s41377-024-01471-9. Light Sci Appl. 2024. PMID: 38834554 Free PMC article.
-
Fishnet mesh of centrin-Sfi1 drives ultrafast calcium-activated contraction of the giant cell Spirostomum ambiguum.bioRxiv [Preprint]. 2024 Nov 8:2024.11.07.622534. doi: 10.1101/2024.11.07.622534. bioRxiv. 2024. PMID: 39574644 Free PMC article. Preprint.
-
Photodegradable polyacrylamide tanglemers enable spatiotemporal control over chain lengthening in high-strength and low-hysteresis hydrogels.J Mater Chem B. 2025 Jan 15;13(3):894-903. doi: 10.1039/d4tb02149e. J Mater Chem B. 2025. PMID: 39648868 Free PMC article.
References
-
- Wilson A. M., et al. , Locomotion dynamics of hunting in wild cheetahs. Nature 498, 185–189 (2013). - PubMed
-
- Yaraghi N. A., et al. , The stomatopod telson: Convergent evolution in the development of a biological shield. Adv. Funct. Mater. 29, 1902238 (2019).
-
- Ha N. S., Lu G., Shu D. W., Yu T. X., Mechanical properties and energy absorption characteristics of tropical fruit durian (Durio zibethinus). J. Mech. Behav. Biomed. Mater. 104, 103603 (2020). - PubMed
-
- Li T. T., Wang H., Huang S. Y., Lou C. W., Lin J. H., Bioinspired foam composites resembling pomelo peel: Structural design and compressive, bursting and cushioning properties. Composites, Part B 172, 290–298 (2019).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
