

Glycoproteomic landscape and structural dynamics of TIM family immune checkpoints enabled by mucinase SmE
- PMID: 37794035
- PMCID: PMC10550946
- DOI: 10.1038/s41467-023-41756-y
Glycoproteomic landscape and structural dynamics of TIM family immune checkpoints enabled by mucinase SmE
Abstract
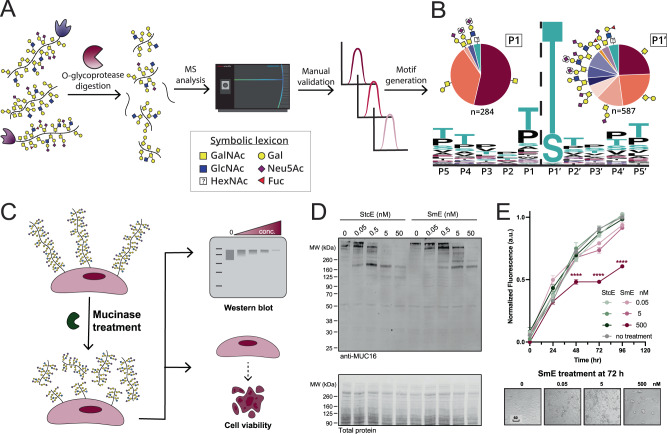
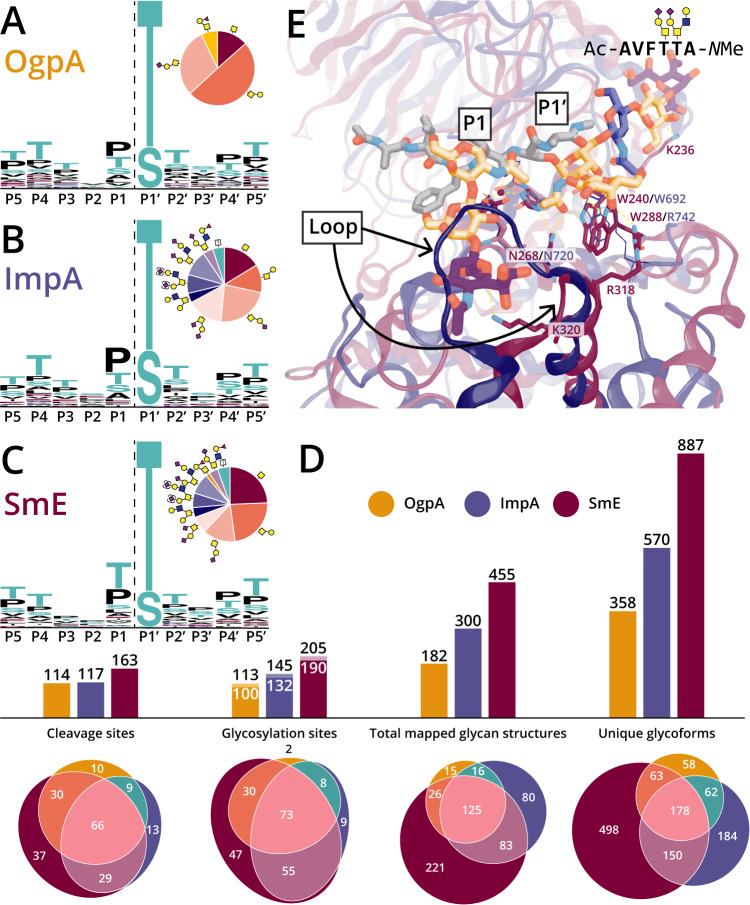
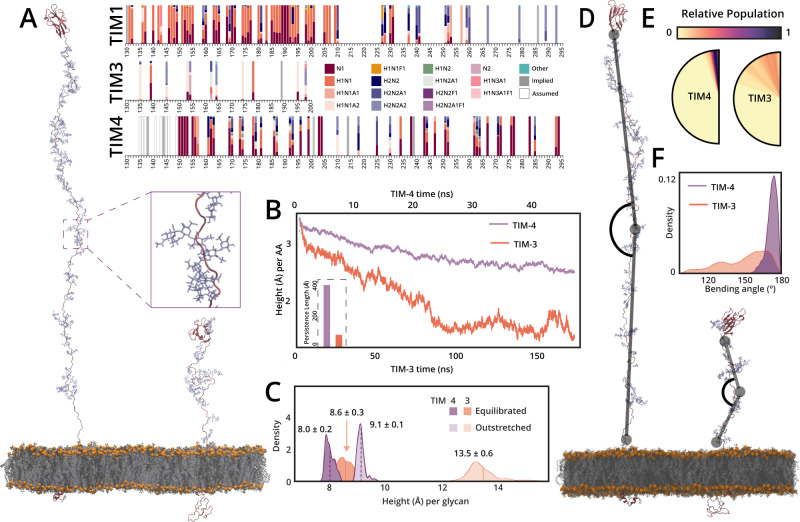
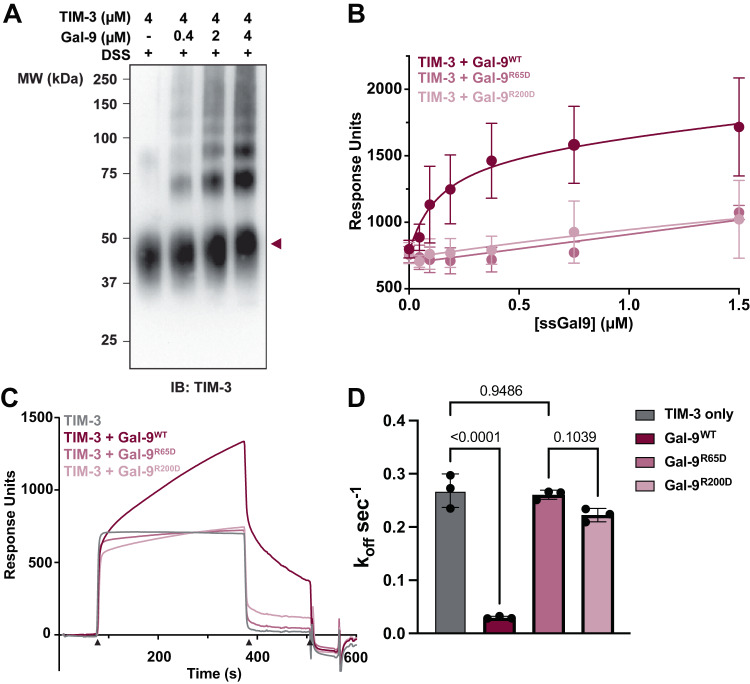
Mucin-domain glycoproteins are densely O-glycosylated and play critical roles in a host of biological functions. In particular, the T cell immunoglobulin and mucin-domain containing family of proteins (TIM-1, -3, -4) decorate immune cells and act as key regulators in cellular immunity. However, their dense O-glycosylation remains enigmatic, primarily due to the challenges associated with studying mucin domains. Here, we demonstrate that the mucinase SmE has a unique ability to cleave at residues bearing very complex glycans. SmE enables improved mass spectrometric analysis of several mucins, including the entire TIM family. With this information in-hand, we perform molecular dynamics (MD) simulations of TIM-3 and -4 to understand how glycosylation affects structural features of these proteins. Finally, we use these models to investigate the functional relevance of glycosylation for TIM-3 function and ligand binding. Overall, we present a powerful workflow to better understand the detailed molecular structures and functions of the mucinome.
© 2023. Springer Nature Limited.
Conflict of interest statement
F.L.K. is a consultant for Protein Evolution, Inc. M.A.H. received consulting fees from Dova Pharmaceuticals, Janssen Pharmaceuticals, and Sonder Capital. C.R.B. is a co-founder and scientific advisory board member of Lycia Therapeutics, Palleon Pharmaceuticals, Enable Bioscience, Redwood Biosciences (a subsidiary of Catalent) OliLux Bio, Grace Science LLC, and InterVenn Biosciences. S.A.M. is a consultant for InterVenn Biosciences and Arkuda Therapeutics. S.A.M., D.J.S., and C.R.B. are coinventors on a Stanford nonprovisional utility patent application that has been filed and is pending in the US (number US20220003777) related to the use of mucinases for mass spectrometry analysis of mucin-domain glycoproteins. The remaining co-authors have no conflicts of interest to disclose.
Figures

Update of
-
Glycoproteomic landscape and structural dynamics of TIM family immune checkpoints enabled by mucinase SmE.bioRxiv [Preprint]. 2023 Feb 3:2023.02.01.526488. doi: 10.1101/2023.02.01.526488. bioRxiv. 2023. Update in: Nat Commun. 2023 Oct 4;14(1):6169. doi: 10.1038/s41467-023-41756-y. PMID: 36778266 Free PMC article. Updated. Preprint.
