

Mitigation of chromosome loss in clinical CRISPR-Cas9-engineered T cells
- PMID: 37794590
- PMCID: PMC10664023
- DOI: 10.1016/j.cell.2023.08.041
Mitigation of chromosome loss in clinical CRISPR-Cas9-engineered T cells
Abstract
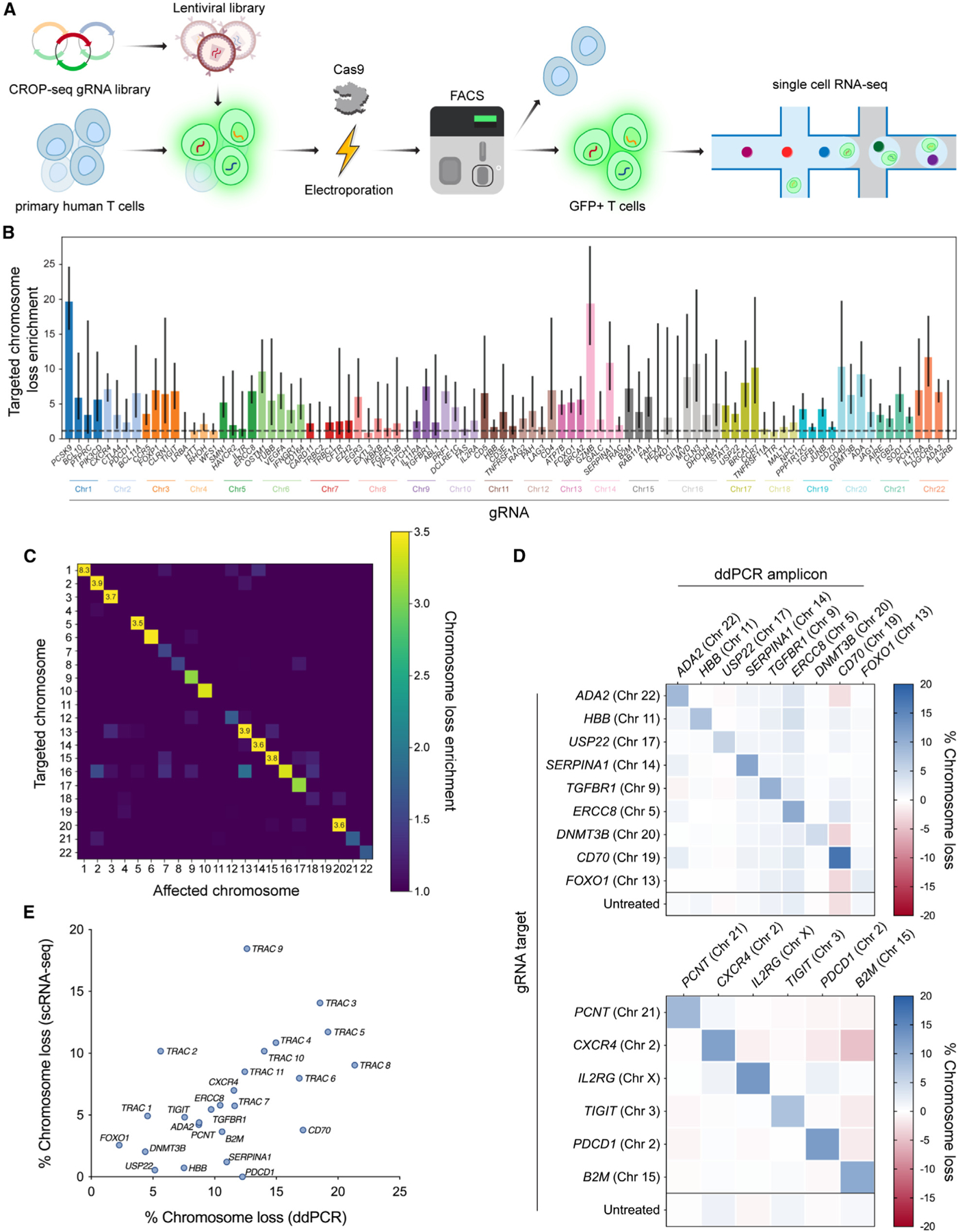
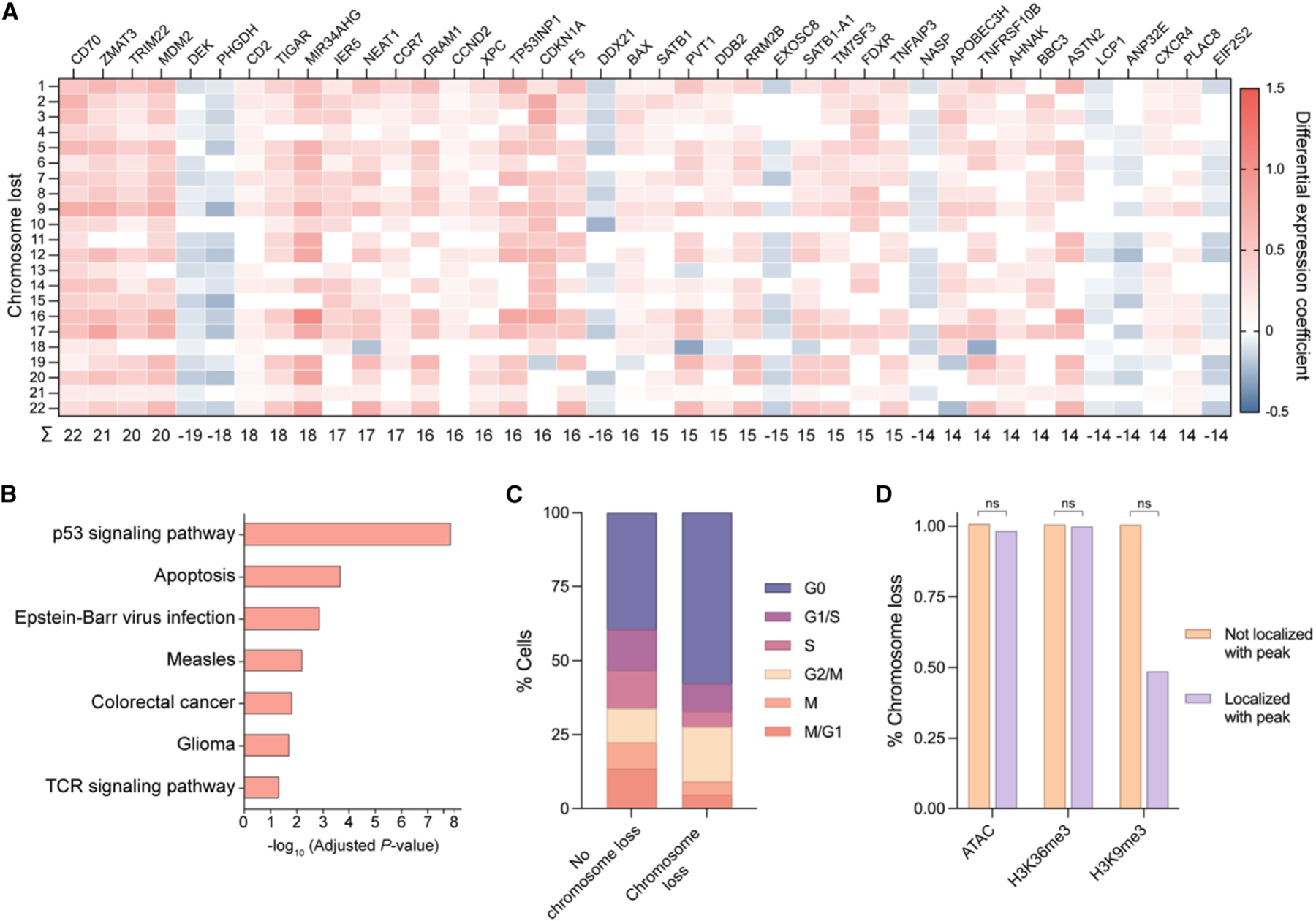
CRISPR-Cas9 genome editing has enabled advanced T cell therapies, but occasional loss of the targeted chromosome remains a safety concern. To investigate whether Cas9-induced chromosome loss is a universal phenomenon and evaluate its clinical significance, we conducted a systematic analysis in primary human T cells. Arrayed and pooled CRISPR screens revealed that chromosome loss was generalizable across the genome and resulted in partial and entire loss of the targeted chromosome, including in preclinical chimeric antigen receptor T cells. T cells with chromosome loss persisted for weeks in culture, implying the potential to interfere with clinical use. A modified cell manufacturing process, employed in our first-in-human clinical trial of Cas9-engineered T cells (NCT03399448), reduced chromosome loss while largely preserving genome editing efficacy. Expression of p53 correlated with protection from chromosome loss observed in this protocol, suggesting both a mechanism and strategy for T cell engineering that mitigates this genotoxicity in the clinic.
Keywords: CAR T cells; CRISPR screen; CRISPR-Cas9; DNA repair; T cells; aneuploidy; chromosome loss; clinical trial; genome editing; immunoengineering.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests C.A.T., J.A.D., and the Regents of the University of California have patents pending or issued related to the use of CRISPR genome editing technologies. R.B. is an employee of BioMarin Pharmaceutical Inc., J.L. is an employee of Altos Labs, and K.R.P. is a co-founder and employee of Cartography Biosciences. A.T.S. is a co-founder of Immunai and Cartography Biosciences. A.T.S. has received research support from Arsenal Biosciences, Allogene Therapeutics, and 10x Genomics. J.H.D.C. is a co-founder of Initial Therapeutics. J.E. is a co-founder of Mnemo Therapeutics, a scientific advisory board member of Cytovia Therapeutics, and a consultant for Casdin Capital, Resolution Therapeutics, IndeeLabs, and Treefrog Therapeutics. J.E. has received research support from Cytovia Therapeutics, Mnemo Therapeutics, and Takeda Pharmaceutical Company. J.A.F. has received research support from Tmunity. C.H.J. and the University of Pennsylvania have patents pending or issued related to the use of gene modification in T cells for adoptive T cell therapy. C.H.J. is a co-founder of Tmunity. H.Y.C. is a co-founder of Accent Therapeutics, Boundless Bio, Cartography Biosciences, and Orbital Therapeutics, and an advisor to 10x Genomics, Arsenal Biosciences, Chroma Medicine, Spring Discovery, and Vida Ventures. C.J.Y. is a co-founder of Survey Genomics, and a scientific advisory board member of Related Sciences and Immunai. C.J.Y. is a consultant for Maze Therapeutics, TReX Bio, ImYoo, and Santa Ana Bio. C.J.Y. has received research support from the Chan Zuckerberg Initiative, Chan Zuckerberg Biohub, Genentech, BioLegend, ScaleBio, and Illumina. J.A.D. is a co-founder of Editas Medicine, Intellia Therapeutics, Caribou Biosciences, Mammoth Biosciences, and Scribe Therapeutics, and a scientific advisory board member of Intellia Therapeutics, Caribou Biosciences, Mammoth Biosciences, Scribe Therapeutics, Vertex Pharmaceuticals, Felix Biosciences, The Column Group, Inari, and Isomorphic Labs. J.A.D. is the Chief Science Advisor at Sixth Street and a Director at Johnson & Johnson, Tempus, and Altos Labs. J.A.D. has sponsored research projects through Apple Tree Partners, Genentech, and Roche.
Figures




Update of
-
Mitigation of chromosome loss in clinical CRISPR-Cas9-engineered T cells.bioRxiv [Preprint]. 2023 Mar 22:2023.03.22.533709. doi: 10.1101/2023.03.22.533709. bioRxiv. 2023. Update in: Cell. 2023 Oct 12;186(21):4567-4582.e20. doi: 10.1016/j.cell.2023.08.041. PMID: 36993359 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
