

Design of hypoxia responsive CRISPR-Cas9 for target gene regulation
- PMID: 37798384
- PMCID: PMC10556097
- DOI: 10.1038/s41598-023-43711-9
Design of hypoxia responsive CRISPR-Cas9 for target gene regulation
Abstract
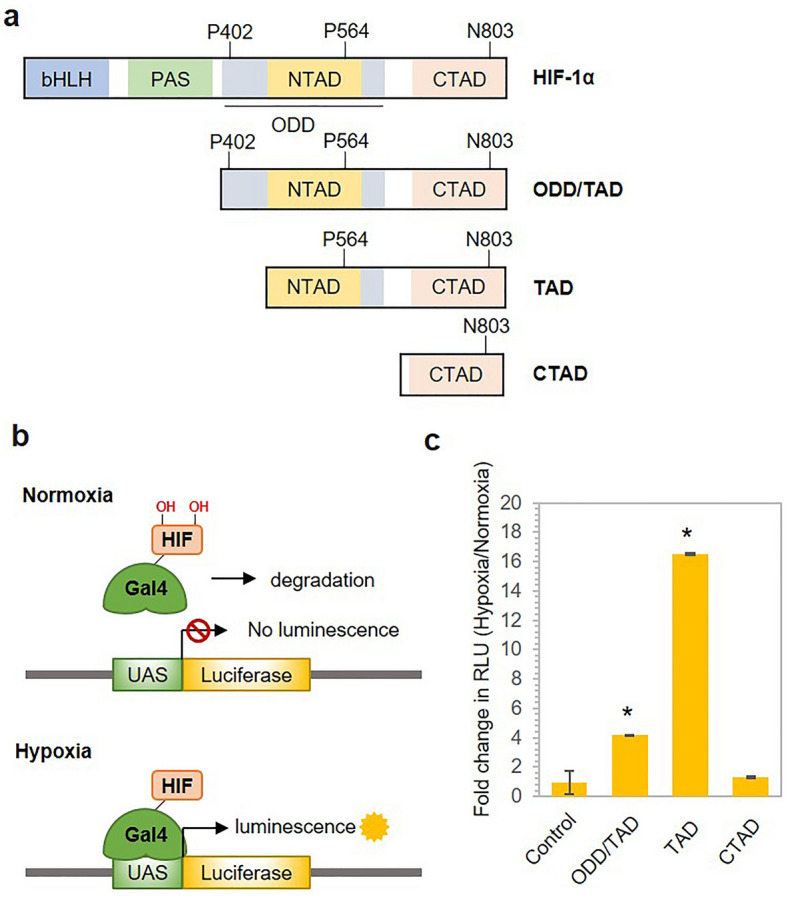
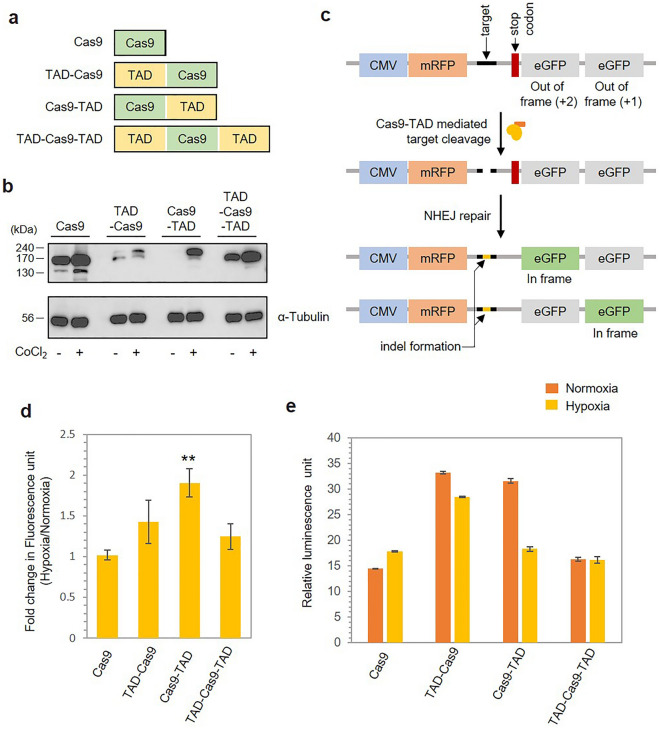
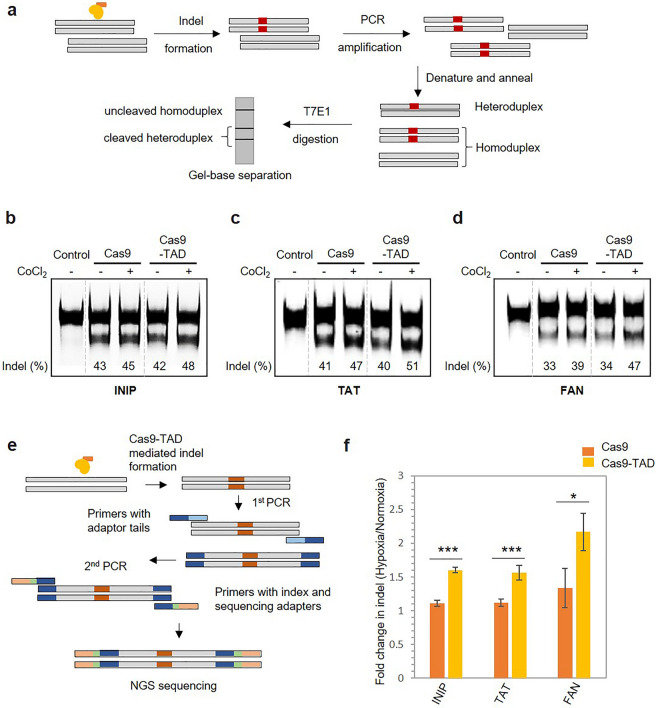
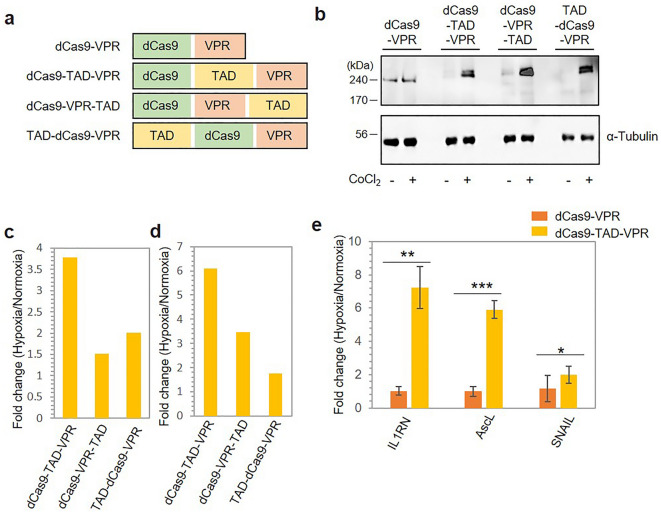
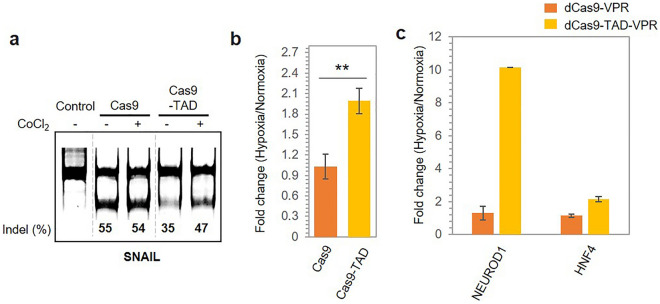
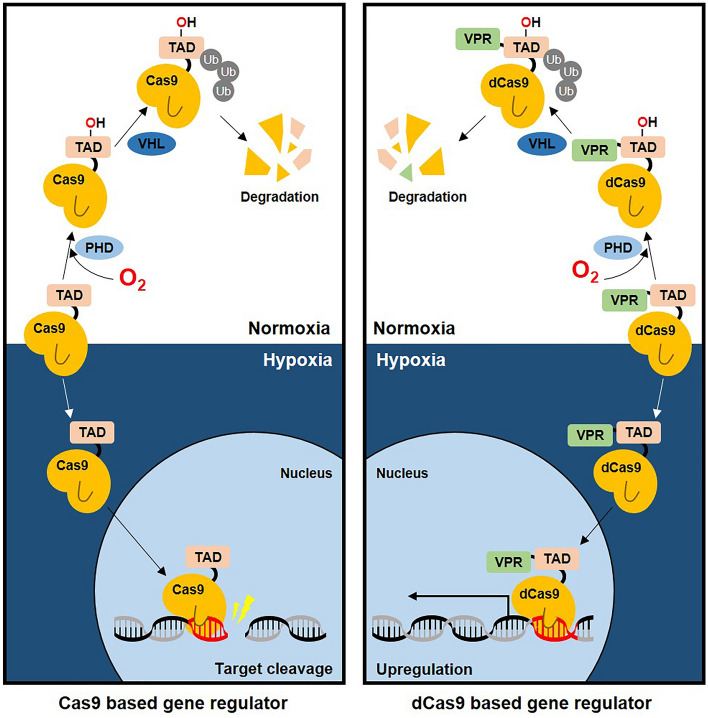
The CRISPR-Cas9 system is a widely used gene-editing tool, offering unprecedented opportunities for treating various diseases. Controlling Cas9/dCas9 activity at specific location and time to avoid undesirable effects is very important. Here, we report a conditionally active CRISPR-Cas9 system that regulates target gene expression upon sensing cellular environmental change. We conjugated the oxygen-sensing transcription activation domain (TAD) of hypoxia-inducing factor (HIF-1α) with the Cas9/dCas9 protein. The Cas9-TAD conjugate significantly increased endogenous target gene cleavage under hypoxic conditions compared with that under normoxic conditions, whereas the dCas9-TAD conjugate upregulated endogenous gene transcription. Furthermore, the conjugate system effectively downregulated the expression of SNAIL, an essential gene in cancer metastasis, and upregulated the expression of the tumour-related genes HNF4 and NEUROD1 under hypoxic conditions. Since hypoxia is closely associated with cancer, the hypoxia-dependent Cas9/dCas9 system is a novel addition to the molecular tool kit that functions in response to cellular signals and has potential application for gene therapeutics.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
CRISPR/Cas9-based epigenome editing: An overview of dCas9-based tools with special emphasis on off-target activity.Methods. 2019 Jul 15;164-165:109-119. doi: 10.1016/j.ymeth.2019.05.003. Epub 2019 May 6. Methods. 2019. PMID: 31071448 Review.
-
Inducible CRISPR-dCas9 Transcriptional Systems for Sensing and Genome Regulation.Chembiochem. 2021 Jun 2;22(11):1894-1900. doi: 10.1002/cbic.202000723. Epub 2021 Mar 3. Chembiochem. 2021. PMID: 33433941
-
dCas9 Tells Tales: Probing Gene Function and Transcription Regulation in Cancer.CRISPR J. 2024 Apr;7(2):73-87. doi: 10.1089/crispr.2023.0078. CRISPR J. 2024. PMID: 38635328 Review.
-
CRISPR/dCas9 Tools: Epigenetic Mechanism and Application in Gene Transcriptional Regulation.Int J Mol Sci. 2023 Oct 3;24(19):14865. doi: 10.3390/ijms241914865. Int J Mol Sci. 2023. PMID: 37834313 Free PMC article. Review.
-
Knockdown of hypoxia-inducible factor-1 alpha by tumor targeted delivery of CRISPR/Cas9 system suppressed the metastasis of pancreatic cancer.J Control Release. 2019 Jun 28;304:204-215. doi: 10.1016/j.jconrel.2019.05.019. Epub 2019 May 14. J Control Release. 2019. PMID: 31100311
Cited by
-
Current Landscape and Future Directions in Cancer Immunotherapy: Therapies, Trials, and Challenges.Cancers (Basel). 2025 Feb 27;17(5):821. doi: 10.3390/cancers17050821. Cancers (Basel). 2025. PMID: 40075668 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
