

Single-nucleus genomics in outbred rats with divergent cocaine addiction-like behaviors reveals changes in amygdala GABAergic inhibition
- PMID: 37798411
- PMCID: PMC10620093
- DOI: 10.1038/s41593-023-01452-y
Single-nucleus genomics in outbred rats with divergent cocaine addiction-like behaviors reveals changes in amygdala GABAergic inhibition
Erratum in
-
Author Correction: Single-nucleus genomics in outbred rats with divergent cocaine addiction-like behaviors reveals changes in amygdala GABAergic inhibition.Nat Neurosci. 2023 Nov;26(11):2035. doi: 10.1038/s41593-023-01489-z. Nat Neurosci. 2023. PMID: 37845545 Free PMC article. No abstract available.
Abstract
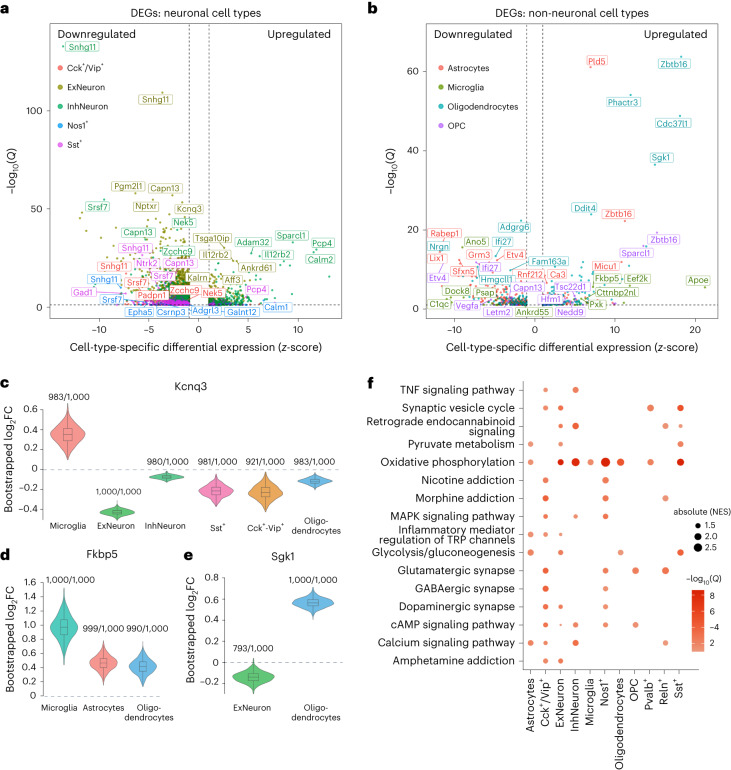
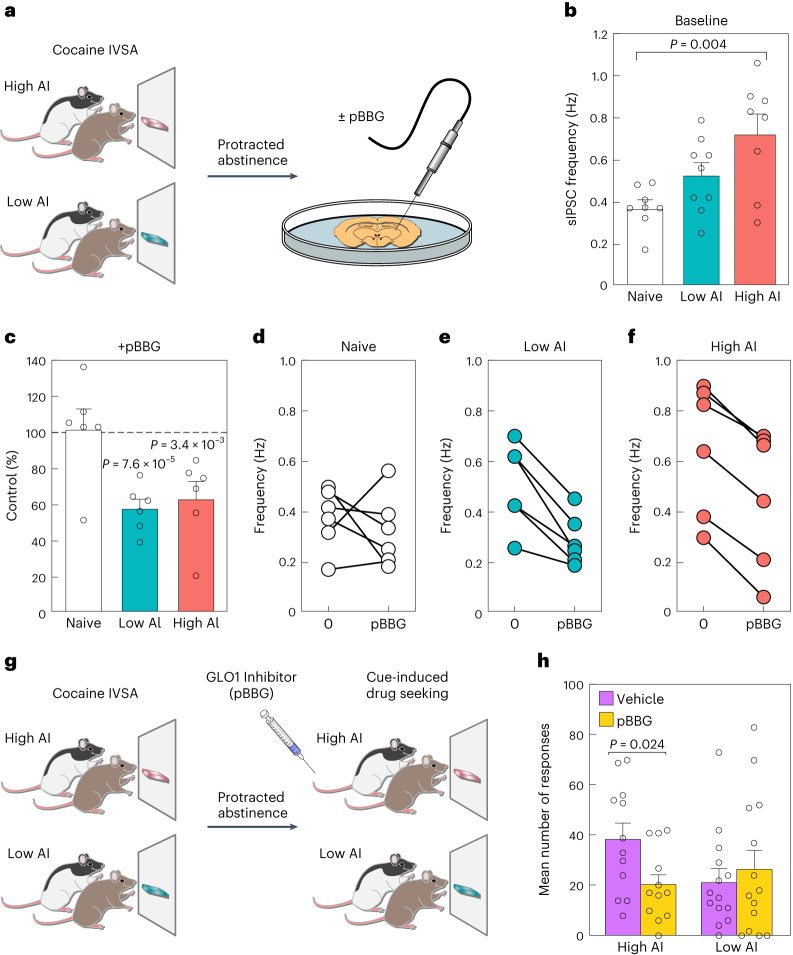
The amygdala processes positive and negative valence and contributes to addiction, but the cell-type-specific gene regulatory programs involved are unknown. We generated an atlas of single-nucleus gene expression and chromatin accessibility in the amygdala of outbred rats with high and low cocaine addiction-like behaviors following prolonged abstinence. Differentially expressed genes between the high and low groups were enriched for energy metabolism across cell types. Rats with high addiction index (AI) showed increased relapse-like behaviors and GABAergic transmission in the amygdala. Both phenotypes were reversed by pharmacological inhibition of the glyoxalase 1 enzyme, which metabolizes methylglyoxal-a GABAA receptor agonist produced by glycolysis. Differences in chromatin accessibility between high and low AI rats implicated pioneer transcription factors in the basic helix-loop-helix, FOX, SOX and activator protein 1 families. We observed opposite regulation of chromatin accessibility across many cell types. Most notably, excitatory neurons had greater accessibility in high AI rats and inhibitory neurons had greater accessibility in low AI rats.
© 2023. The Author(s).
Conflict of interest statement
A.A.P. holds a patent related to the use of GLO1 inhibitors (US20160038559, active). The inventors of this patent are A. Palmer and M. Distler. All other authors declare no competing interests.
Figures



References
-
- Buffalari, D. M. & See, R. E. in Behavioral Neuroscience ofDrug Addiction (eds. Self, D. W. & Staley Gottschalk, J. K.) (Springer, 2010); 10.1007/7854_2009_18
-
- Koob, G. F. in Anhedonia: Preclinical, Translational, and Clinical Integration (ed. Pizzagalli, D. A.) (Springer, 2022); 10.1007/7854_2021_288
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
