

The heme-responsive PrrH sRNA regulates Pseudomonas aeruginosa pyochelin gene expression
- PMID: 37800921
- PMCID: PMC10597452
- DOI: 10.1128/msphere.00392-23
The heme-responsive PrrH sRNA regulates Pseudomonas aeruginosa pyochelin gene expression
Abstract
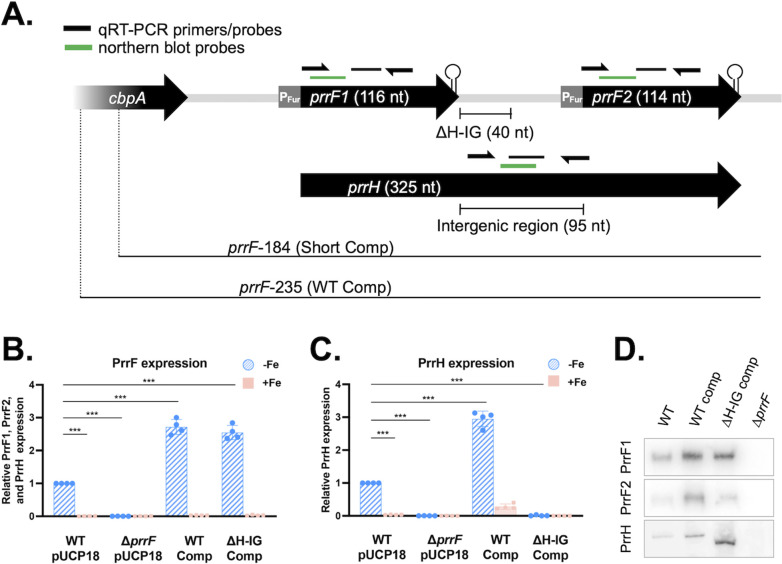
Pseudomonas aeruginosa is an opportunistic pathogen that requires iron for growth and virulence, yet this nutrient is sequestered by the innate immune system during infection. When iron is limiting, P. aeruginosa expresses the PrrF1 and PrrF2 small RNAs (sRNAs), which post-transcriptionally repress expression of nonessential iron-containing proteins, thus sparing this nutrient for more critical processes. The genes for the PrrF1 and PrrF2 sRNAs are arranged in tandem on the chromosome, allowing for the transcription of a longer heme-responsive sRNA, termed PrrH. While the functions of PrrF1 and PrrF2 have been extensively studied, the role of PrrH in P. aeruginosa physiology and virulence is not well understood. In this study, we performed transcriptomic and proteomic studies to identify the PrrH regulon. In shaking cultures, the pyochelin synthesis proteins were increased in two distinct prrH mutants compared to the wild type, while the mRNAs for these proteins were not affected by the prrH mutation. We identified complementarity between the PrrH sRNA and the sequence upstream of the pchE mRNA, suggesting the potential for PrrH to directly regulate the expression of genes for pyochelin synthesis. We further showed that pchE mRNA levels were increased in the prrH mutants when grown in static but not shaking conditions. Moreover, we discovered that controlling for the presence of light was critical for examining the impact of PrrH on pchE expression. As such, our study reports on the first likely target of the PrrH sRNA and highlights key environmental variables that will allow for future characterization of PrrH function. IMPORTANCE In the human host, iron is predominantly in the form of heme, which Pseudomonas aeruginosa can acquire as an iron source during infection. We previously showed that the iron-responsive PrrF small RNAs (sRNAs) are critical for mediating iron homeostasis during P. aeruginosa infection; however, the function of the heme-responsive PrrH sRNA remains unclear. In this study, we identified genes for pyochelin siderophore biosynthesis, which mediates uptake of inorganic iron, as a novel target of PrrH regulation. This study therefore highlights a novel relationship between heme availability and siderophore biosynthesis in P. aeruginosa.
Keywords: PrrF; PrrH; Pseudomonas aeruginosa; heme; iron; pyochelin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Update of
-
The heme-responsive PrrH sRNA regulates Pseudomonas aeruginosa pyochelin gene expression.bioRxiv [Preprint]. 2023 Jan 20:2023.01.19.524833. doi: 10.1101/2023.01.19.524833. bioRxiv. 2023. Update in: mSphere. 2023 Oct 24;8(5):e0039223. doi: 10.1128/msphere.00392-23. PMID: 36712080 Free PMC article. Updated. Preprint.
Similar articles
-
The heme-responsive PrrH sRNA regulates Pseudomonas aeruginosa pyochelin gene expression.bioRxiv [Preprint]. 2023 Jan 20:2023.01.19.524833. doi: 10.1101/2023.01.19.524833. bioRxiv. 2023. Update in: mSphere. 2023 Oct 24;8(5):e0039223. doi: 10.1128/msphere.00392-23. PMID: 36712080 Free PMC article. Updated. Preprint.
-
The prrF-encoded small regulatory RNAs are required for iron homeostasis and virulence of Pseudomonas aeruginosa.Infect Immun. 2015 Mar;83(3):863-75. doi: 10.1128/IAI.02707-14. Epub 2014 Dec 15. Infect Immun. 2015. PMID: 25510881 Free PMC article.
-
The Pseudomonas aeruginosa PrrF Small RNAs Regulate Iron Homeostasis during Acute Murine Lung Infection.Infect Immun. 2017 Apr 21;85(5):e00764-16. doi: 10.1128/IAI.00764-16. Print 2017 May. Infect Immun. 2017. PMID: 28289146 Free PMC article.
-
How we learnt about iron acquisition in Pseudomonas aeruginosa: a series of very fortunate events.Biometals. 2007 Jun;20(3-4):587-601. doi: 10.1007/s10534-006-9067-2. Epub 2006 Dec 22. Biometals. 2007. PMID: 17186376 Review.
-
Pseudomonas aeruginosa virulence attenuation by inhibiting siderophore functions.Appl Microbiol Biotechnol. 2023 Feb;107(4):1019-1038. doi: 10.1007/s00253-022-12347-6. Epub 2023 Jan 12. Appl Microbiol Biotechnol. 2023. PMID: 36633626 Review.
Cited by
-
Low leucine levels in the blood enhance the pathogenicity of neonatal meningitis-causing Escherichia coli.Nat Commun. 2025 Mar 12;16(1):2466. doi: 10.1038/s41467-025-57850-2. Nat Commun. 2025. PMID: 40075077 Free PMC article.
-
Pseudomonas aeruginosa heme metabolites biliverdin IXβ and IXδ are integral to lifestyle adaptations associated with chronic infection.mBio. 2024 Mar 13;15(3):e0276323. doi: 10.1128/mbio.02763-23. Epub 2024 Feb 6. mBio. 2024. PMID: 38319089 Free PMC article.
-
Calprotectin elicits aberrant iron starvation responses in Pseudomonas aeruginosa under anaerobic conditions.J Bacteriol. 2025 Apr 17;207(4):e0002925. doi: 10.1128/jb.00029-25. Epub 2025 Mar 26. J Bacteriol. 2025. PMID: 40135923 Free PMC article.
-
The Pseudomonas aeruginosa PhuS proximal ligand His-209 triggers a conformational switch in function from DNA binding to heme transfer.J Biol Chem. 2025 May;301(5):108440. doi: 10.1016/j.jbc.2025.108440. Epub 2025 Mar 22. J Biol Chem. 2025. PMID: 40122173 Free PMC article.
-
The Quorum Sensing Regulated sRNA Lrs1 Is Involved in the Adaptation to Low Iron in Pseudomonas aeruginosa.Environ Microbiol Rep. 2025 Apr;17(2):e70090. doi: 10.1111/1758-2229.70090. Environ Microbiol Rep. 2025. PMID: 40150866 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
