

Neuraminidase inhibition improves endothelial function in diabetic mice
- PMID: 37801046
- PMCID: PMC10908409
- DOI: 10.1152/ajpheart.00337.2023
Neuraminidase inhibition improves endothelial function in diabetic mice
Abstract
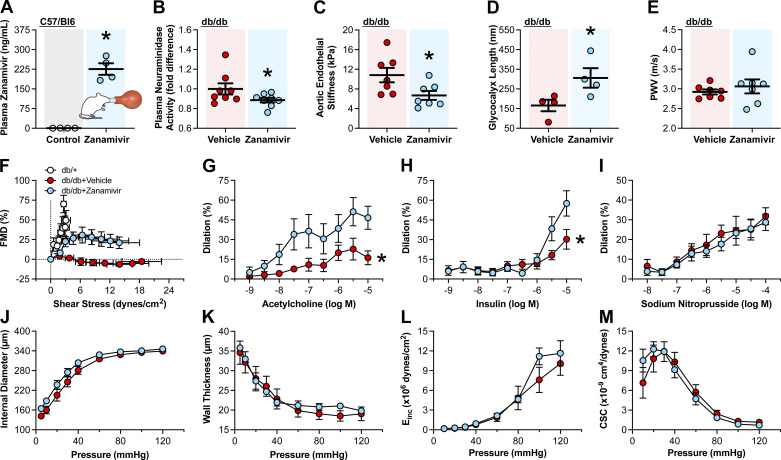
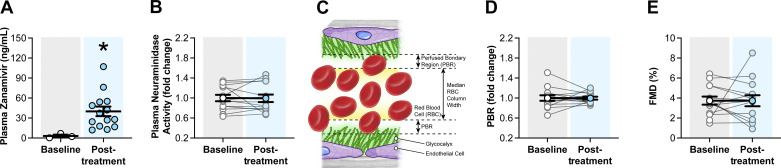
Neuraminidases cleave sialic acids from glycocalyx structures and plasma neuraminidase activity is elevated in type 2 diabetes (T2D). Therefore, we hypothesize circulating neuraminidase degrades the endothelial glycocalyx and diminishes flow-mediated dilation (FMD), whereas its inhibition restores shear mechanosensation and endothelial function in T2D settings. We found that compared with controls, subjects with T2D have higher plasma neuraminidase activity, reduced plasma nitrite concentrations, and diminished FMD. Ex vivo and in vivo neuraminidase exposure diminished FMD and reduced endothelial glycocalyx presence in mouse arteries. In cultured endothelial cells, neuraminidase reduced glycocalyx coverage. Inhalation of the neuraminidase inhibitor, zanamivir, reduced plasma neuraminidase activity, enhanced endothelial glycocalyx length, and improved FMD in diabetic mice. In humans, a single-arm trial (NCT04867707) of zanamivir inhalation did not reduce plasma neuraminidase activity, improved glycocalyx length, or enhanced FMD. Although zanamivir plasma concentrations in mice reached 225.8 ± 22.0 ng/mL, in humans were only 40.0 ± 7.2 ng/mL. These results highlight the potential of neuraminidase inhibition for ameliorating endothelial dysfunction in T2D and suggest the current Food and Drug Administration-approved inhaled dosage of zanamivir is insufficient to achieve desired outcomes in humans.NEW & NOTEWORTHY This work identifies neuraminidase as a key mediator of endothelial dysfunction in type 2 diabetes that may serve as a biomarker for impaired endothelial function and predictive of development and progression of cardiovascular pathologies associated with type 2 diabetes (T2D). Data show that intervention with the neuraminidase inhibitor zanamivir at effective plasma concentrations may represent a novel pharmacological strategy for restoring the glycocalyx and ameliorating endothelial dysfunction.
Keywords: flow-mediated dilation; glycocalyx; type 2 diabetes; zanamivir.
Conflict of interest statement
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Pending US patent Application, Serial No. 16/754,814. Entitled: Neuraminidase Inhibition to Improve Glycocalyx Volume and Function to Ameliorate Cardiovascular Diseases in Pathologies Associated with Glycocalyx Damage (filed, 9 April 2020; Ref. No. 17UMC103).
Figures





References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical