

XPO1 Enables Adaptive Regulation of mRNA Export Required for Genotoxic Stress Tolerance in Cancer Cells
- PMID: 37801604
- PMCID: PMC10758694
- DOI: 10.1158/0008-5472.CAN-23-1992
XPO1 Enables Adaptive Regulation of mRNA Export Required for Genotoxic Stress Tolerance in Cancer Cells
Abstract
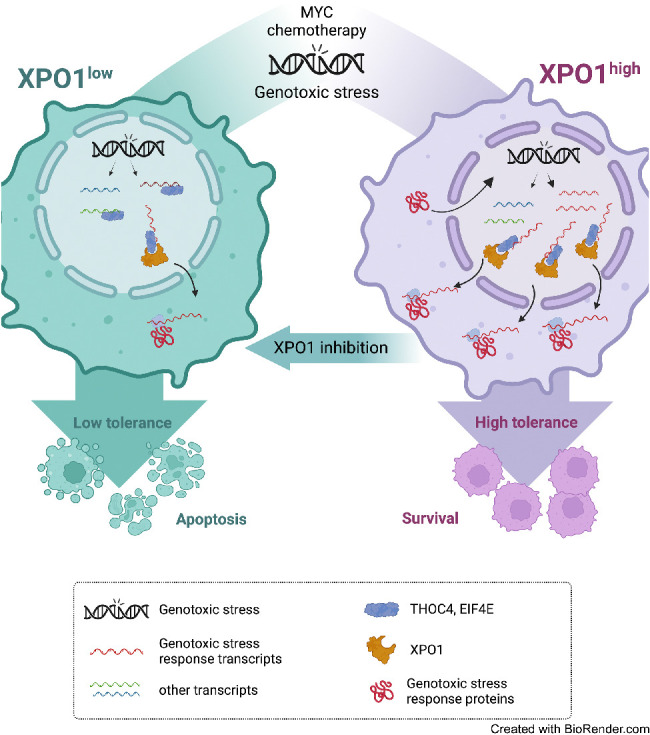
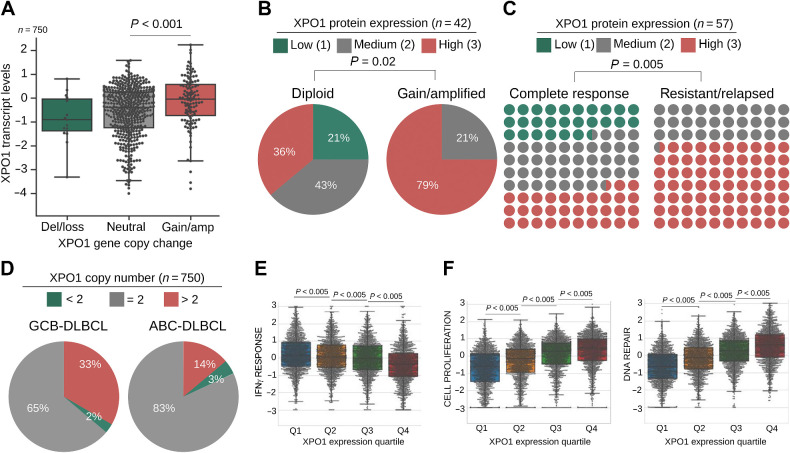
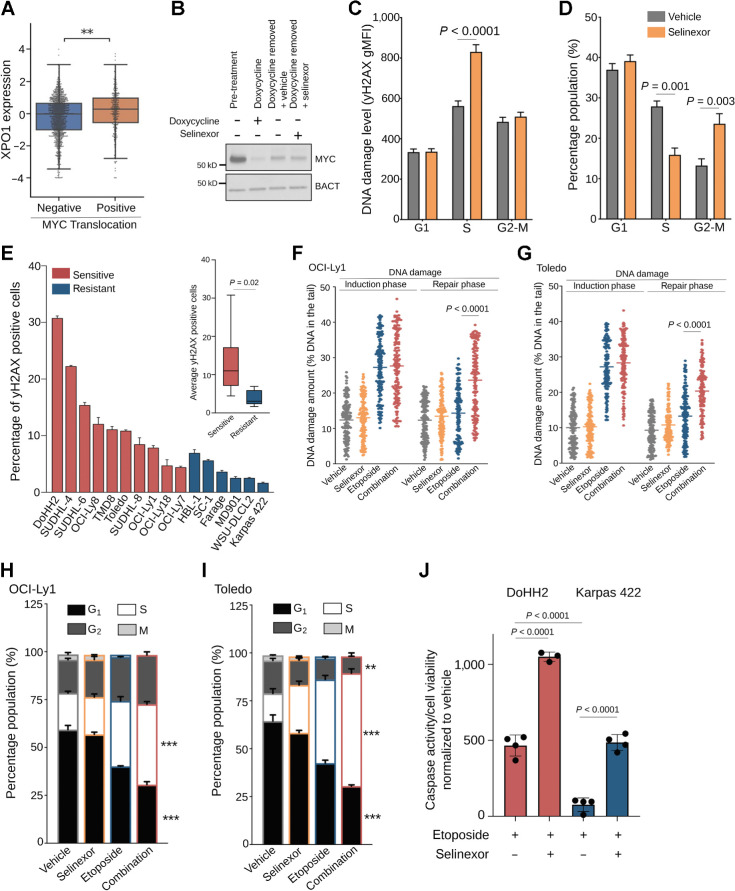
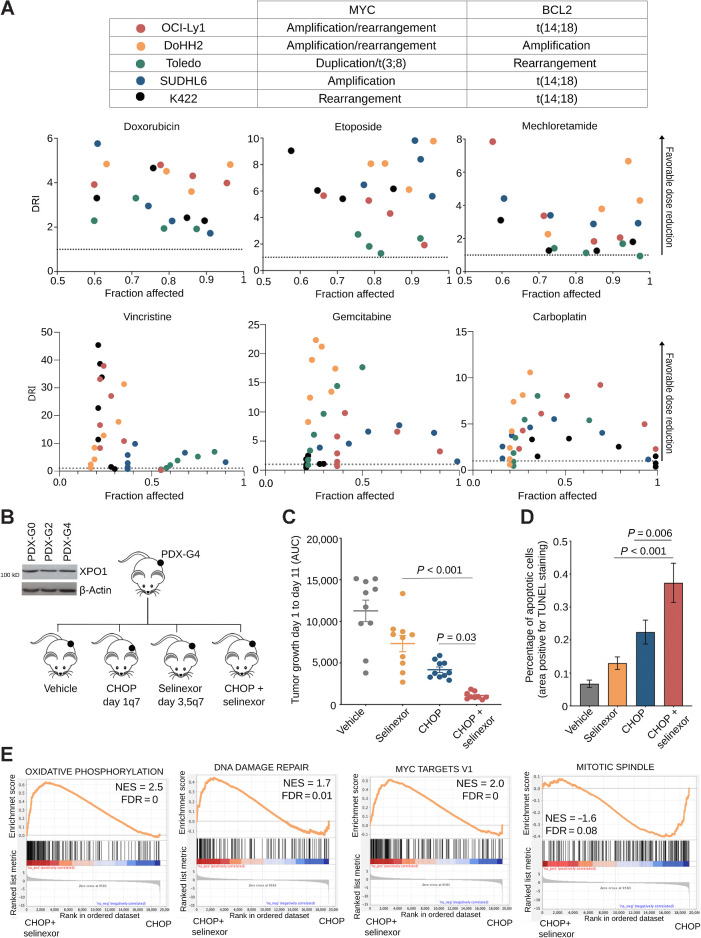
Exportin-1 (XPO1), the main soluble nuclear export receptor in eukaryotic cells, is frequently overexpressed in diffuse large B-cell lymphoma (DLBCL). A selective XPO1 inhibitor, selinexor, received approval as single agent for relapsed or refractory (R/R) DLBCL. Elucidating the mechanisms by which XPO1 overexpression supports cancer cells could facilitate further clinical development of XPO1 inhibitors. We uncovered here that XPO1 overexpression increases tolerance to genotoxic stress, leading to a poor response to chemoimmunotherapy. Upon DNA damage induced by MYC expression or exogenous compounds, XPO1 bound and exported EIF4E and THOC4 carrying DNA damage repair mRNAs, thereby increasing synthesis of DNA damage repair proteins under conditions of increased turnover. Consequently, XPO1 inhibition decreased the capacity of lymphoma cells to repair DNA damage and ultimately resulted in increased cytotoxicity. In a phase I clinical trial conducted in R/R DLBCL, the combination of selinexor with second-line chemoimmunotherapy was tolerated with early indication of efficacy. Overall, this study reveals that XPO1 overexpression plays a critical role in the increased tolerance of cancer cells to DNA damage while providing new insights to optimize the clinical development of XPO1 inhibitors.
Significance: XPO1 regulates the dynamic ribonucleoprotein nuclear export in response to genotoxic stress to support tolerance and can be targeted to enhance the sensitivity of cancer cells to endogenous and exogenous DNA damage. See related commentary by Knittel and Reinhardt, p. 3.
©2023 The Authors; Published by the American Association for Cancer Research.
Figures
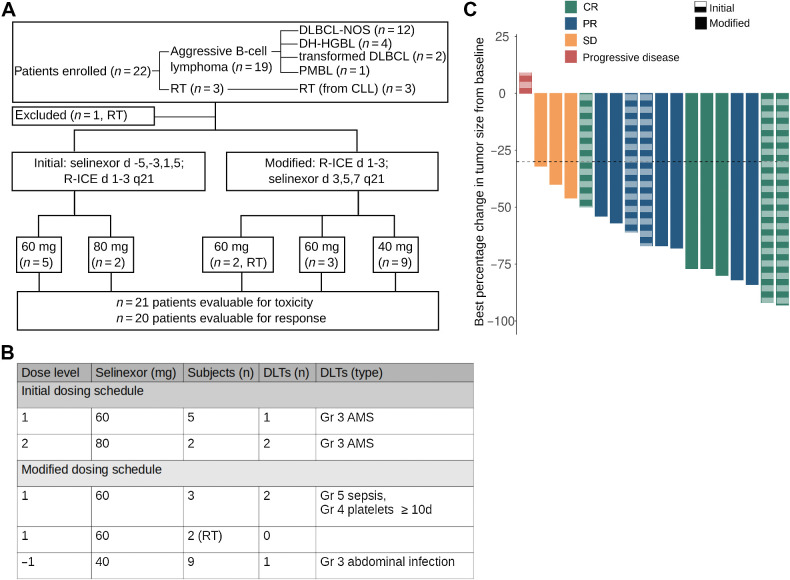
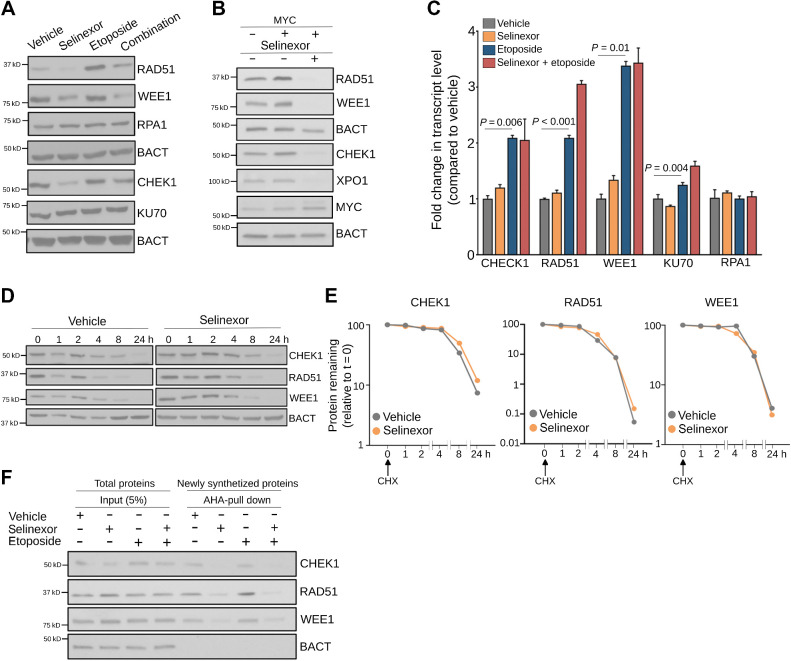
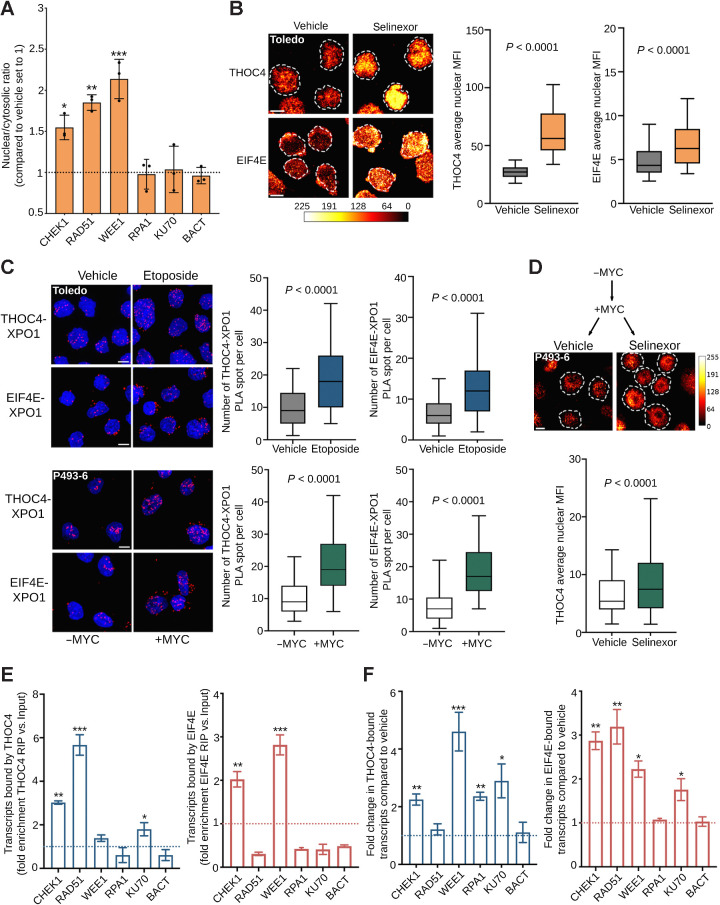
Comment in
-
XPO1-Mediated mRNA Export of Genome Maintenance Regulators Drives Chemotherapy Resistance in Aggressive Lymphoma.Cancer Res. 2024 Jan 2;84(1):3-5. doi: 10.1158/0008-5472.CAN-23-2966. Cancer Res. 2024. PMID: 37902414
References
-
- Zimmerli CE, Allegretti M, Rantos V, Goetz SK, Obarska-Kosinska A, Zagoriy I, et al. Nuclear pores dilate and constrict in cellulo. Science 2021;374:eabd9776. - PubMed
-
- Strambio-De-Castillia C, Niepel M, Rout MP. The nuclear pore complex: bridging nuclear transport and gene regulation. Nat Rev Mol Cell Biol 2010;11:490–501. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
