

Comprehensive quantification of metabolic flux during acute cold stress in mice
- PMID: 37802078
- PMCID: PMC10840821
- DOI: 10.1016/j.cmet.2023.09.002
Comprehensive quantification of metabolic flux during acute cold stress in mice
Abstract
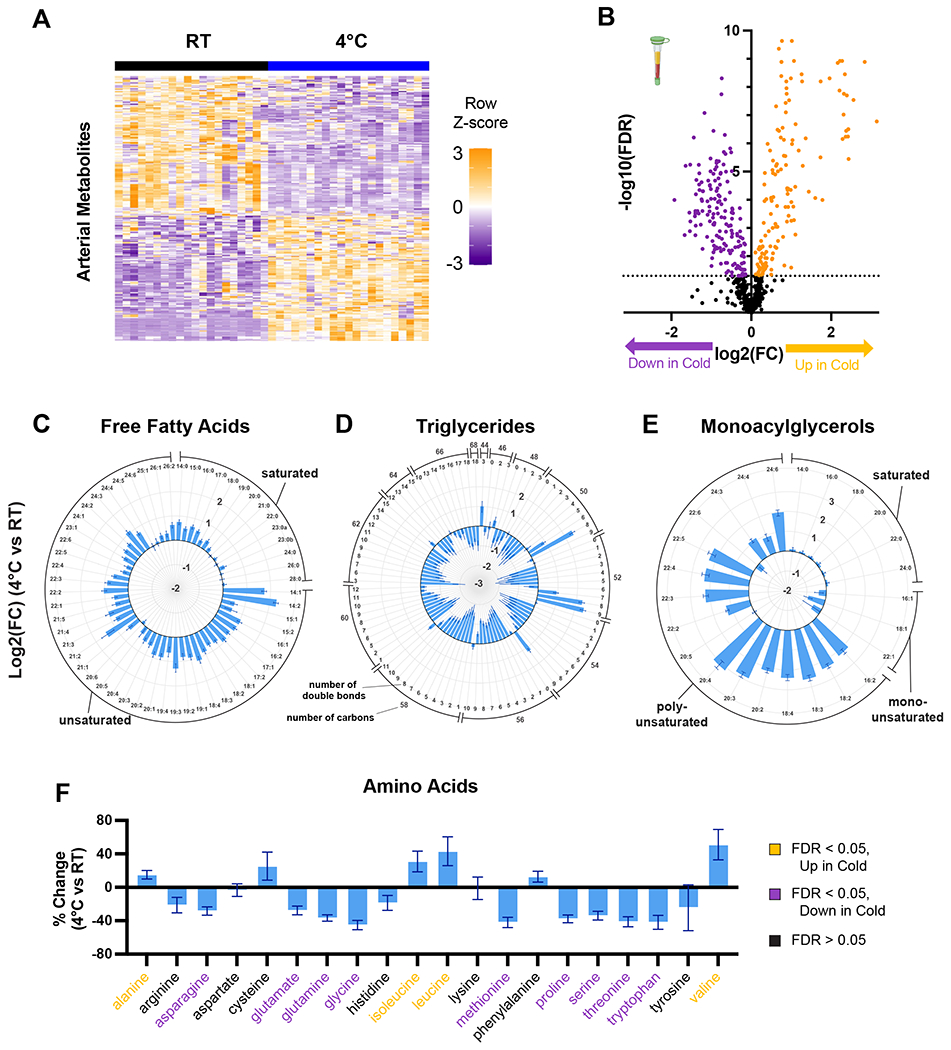
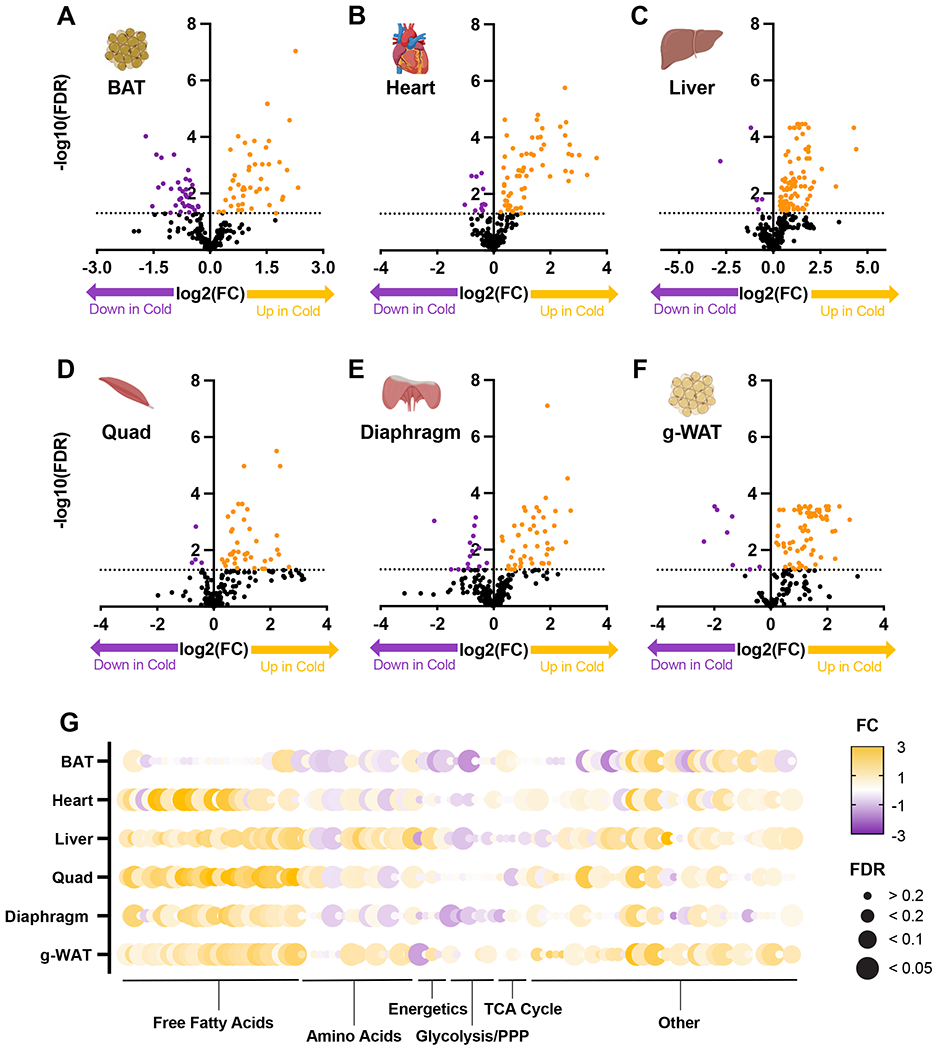
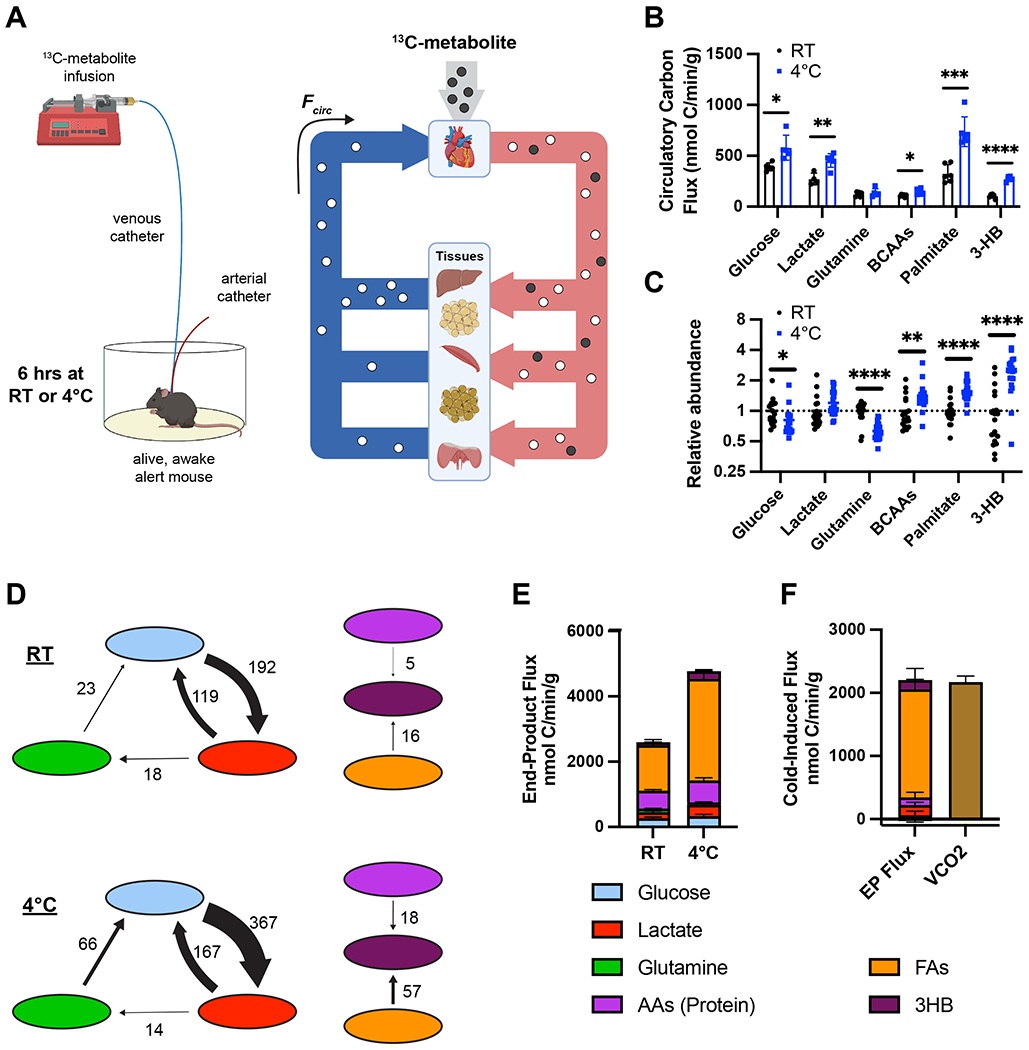
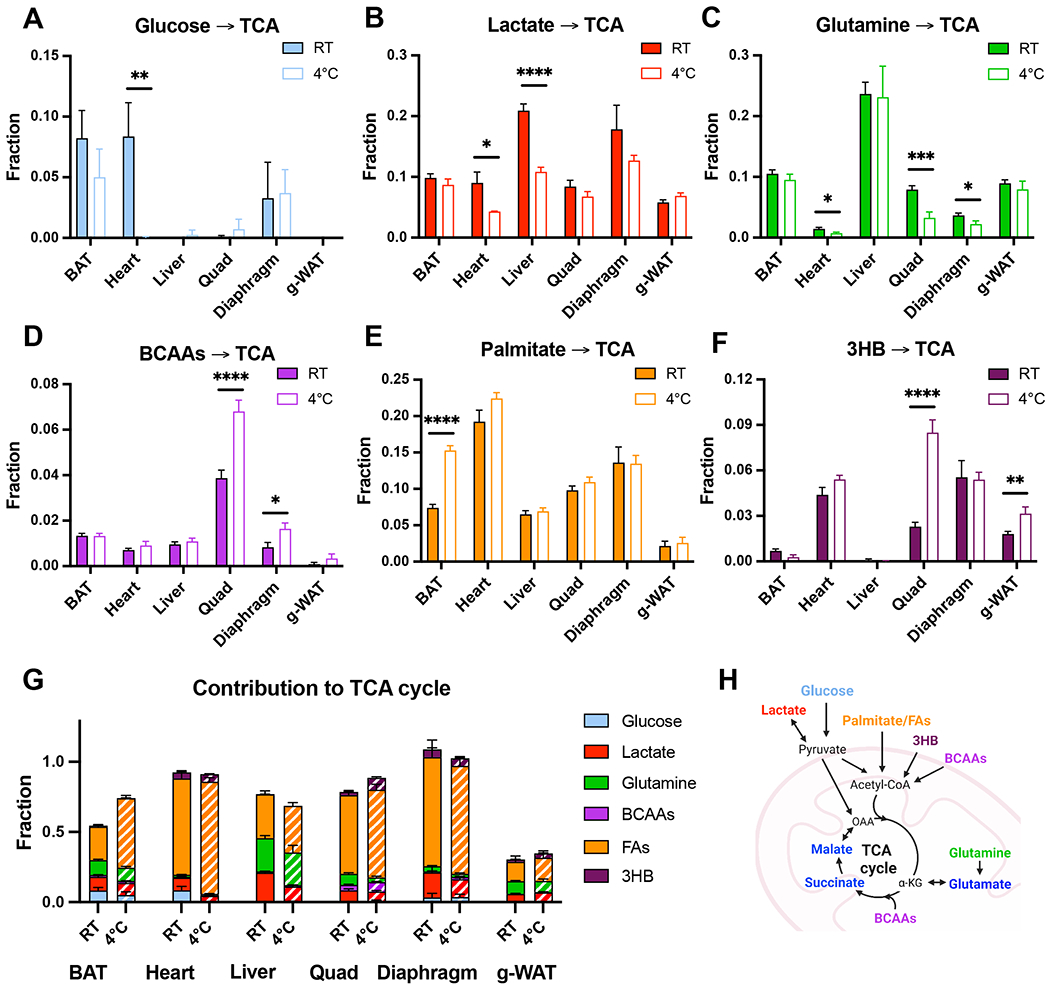
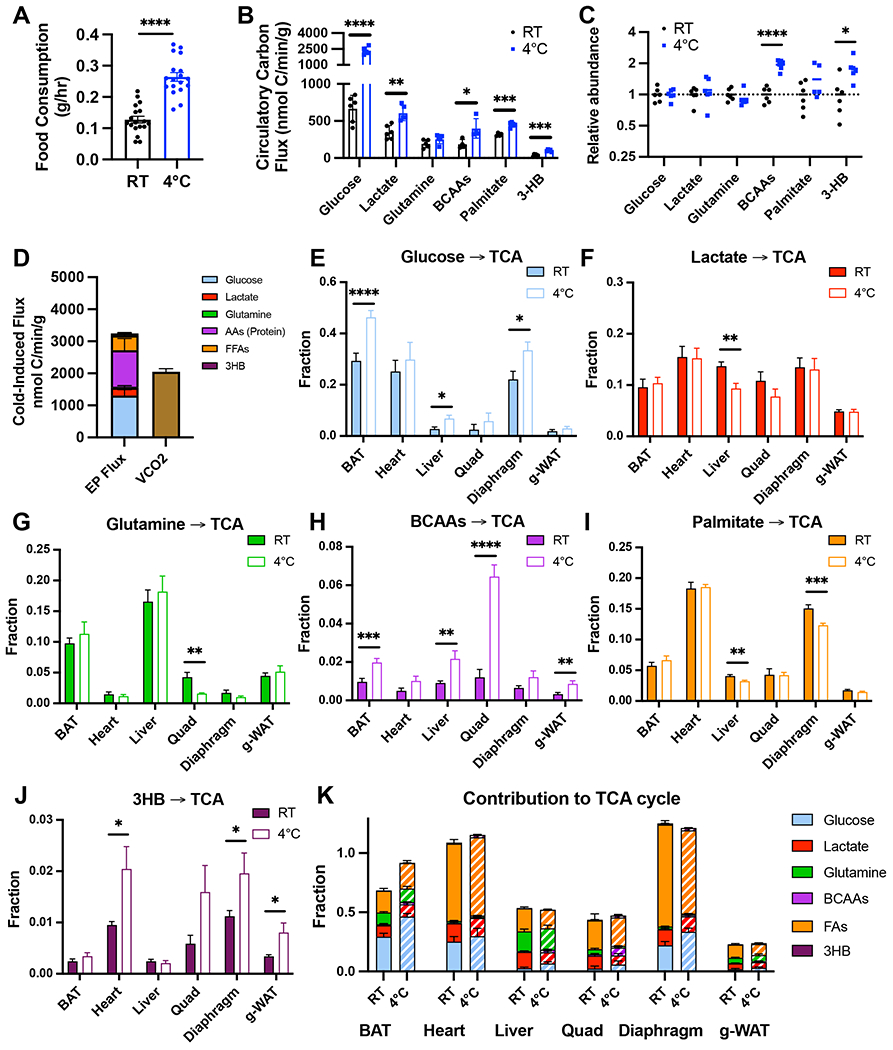
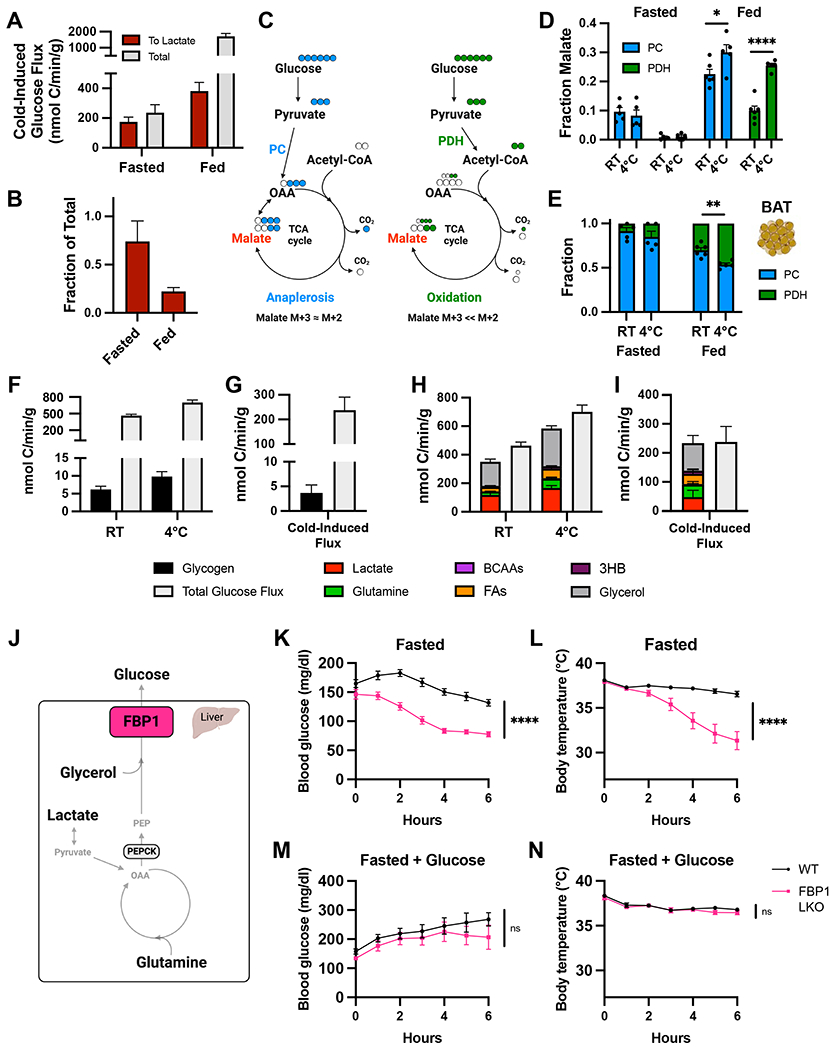
Cold-induced thermogenesis (CIT) is widely studied as a potential avenue to treat obesity, but a thorough understanding of the metabolic changes driving CIT is lacking. Here, we present a comprehensive and quantitative analysis of the metabolic response to acute cold exposure, leveraging metabolomic profiling and minimally perturbative isotope tracing studies in unanesthetized mice. During cold exposure, brown adipose tissue (BAT) primarily fueled the tricarboxylic acid (TCA) cycle with fat in fasted mice and glucose in fed mice, underscoring BAT's metabolic flexibility. BAT minimally used branched-chain amino acids or ketones, which were instead avidly consumed by muscle during cold exposure. Surprisingly, isotopic labeling analyses revealed that BAT uses glucose largely for TCA anaplerosis via pyruvate carboxylation. Finally, we find that cold-induced hepatic gluconeogenesis is critical for CIT during fasting, demonstrating a key functional role for glucose metabolism. Together, these findings provide a detailed map of the metabolic rewiring driving acute CIT.
Keywords: FBP1; brown adipose tissue; cold exposure; flux; gluconeogenesis; glucose; metabolomics; pyruvate carboxylase; thermogenesis.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Commission IT (2003). Glossary of terms for thermal physiology. J. Therm. Biol 28, 75–106. 10.1152/jappl.1973.35.6.941. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
