

Activity-dependent organization of prefrontal hub-networks for associative learning and signal transformation
- PMID: 37803014
- PMCID: PMC10558457
- DOI: 10.1038/s41467-023-41547-5
Activity-dependent organization of prefrontal hub-networks for associative learning and signal transformation
Abstract
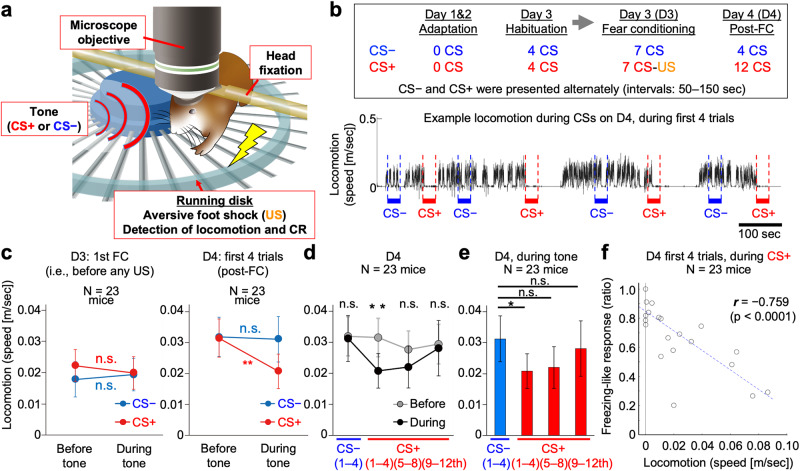
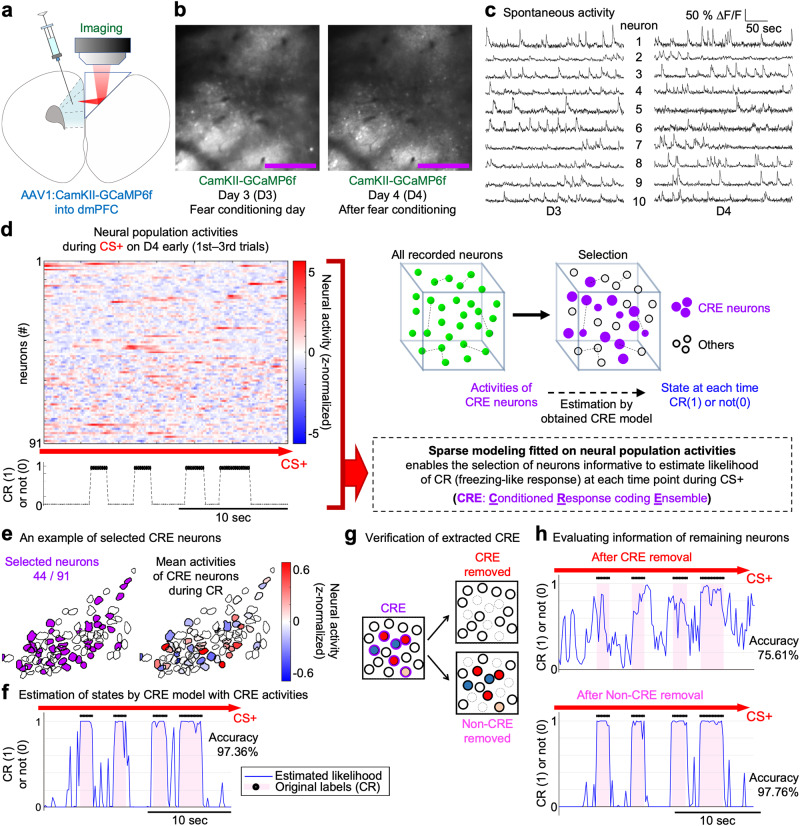
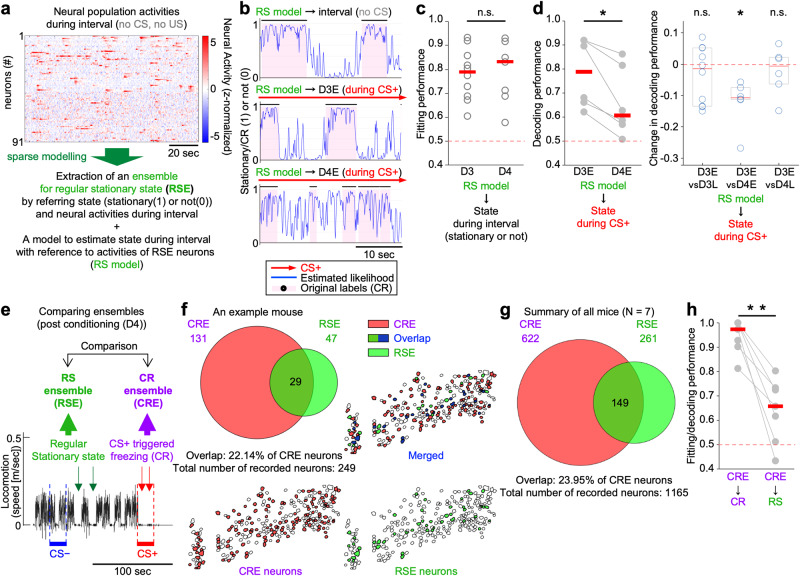
Associative learning is crucial for adapting to environmental changes. Interactions among neuronal populations involving the dorso-medial prefrontal cortex (dmPFC) are proposed to regulate associative learning, but how these neuronal populations store and process information about the association remains unclear. Here we developed a pipeline for longitudinal two-photon imaging and computational dissection of neural population activities in male mouse dmPFC during fear-conditioning procedures, enabling us to detect learning-dependent changes in the dmPFC network topology. Using regularized regression methods and graphical modeling, we found that fear conditioning drove dmPFC reorganization to generate a neuronal ensemble encoding conditioned responses (CR) characterized by enhanced internal coactivity, functional connectivity, and association with conditioned stimuli (CS). Importantly, neurons strongly responding to unconditioned stimuli during conditioning subsequently became hubs of this novel associative network for the CS-to-CR transformation. Altogether, we demonstrate learning-dependent dynamic modulation of population coding structured on the activity-dependent formation of the hub network within the dmPFC.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Prefrontal neuronal assemblies temporally control fear behaviour.Nature. 2016 Jul 21;535(7612):420-4. doi: 10.1038/nature18630. Epub 2016 Jul 13. Nature. 2016. PMID: 27409809
-
The role of prefrontal cortex in predictive fear learning.Behav Neurosci. 2010 Oct;124(5):574-86. doi: 10.1037/a0020739. Behav Neurosci. 2010. PMID: 20939658
-
Learning pain-related fear: neural mechanisms mediating rapid differential conditioning, extinction and reinstatement processes in human visceral pain.Neurobiol Learn Mem. 2014 Dec;116:36-45. doi: 10.1016/j.nlm.2014.08.003. Epub 2014 Aug 13. Neurobiol Learn Mem. 2014. PMID: 25128878
-
Learning-induced intrinsic and synaptic plasticity in the rodent medial prefrontal cortex.Neurobiol Learn Mem. 2020 Mar;169:107117. doi: 10.1016/j.nlm.2019.107117. Epub 2019 Nov 23. Neurobiol Learn Mem. 2020. PMID: 31765801 Free PMC article. Review.
-
Functional basis of associative learning and its relationships with long-term potentiation evoked in the involved neural circuits: Lessons from studies in behaving mammals.Neurobiol Learn Mem. 2015 Oct;124:3-18. doi: 10.1016/j.nlm.2015.04.006. Epub 2015 Apr 25. Neurobiol Learn Mem. 2015. PMID: 25916668 Review.
Cited by
-
Surgery-induced neuroinflammatory transcriptional programs in medial prefrontal cortex of mice during early phase of perioperative neurocognitive disorders.PeerJ. 2024 Dec 3;12:e18664. doi: 10.7717/peerj.18664. eCollection 2024. PeerJ. 2024. PMID: 39650559 Free PMC article.
-
Large-scale cranial window for in vivo mouse brain imaging utilizing fluoropolymer nanosheet and light-curable resin.Commun Biol. 2024 Mar 4;7(1):232. doi: 10.1038/s42003-024-05865-8. Commun Biol. 2024. PMID: 38438546 Free PMC article.
-
Minimally invasive, wide-field two-photon imaging of the brainstem at cellular resolution.Cell Rep Methods. 2025 Apr 21;5(4):101010. doi: 10.1016/j.crmeth.2025.101010. Epub 2025 Apr 4. Cell Rep Methods. 2025. PMID: 40187350 Free PMC article.
-
Prefrontal encoding of an internal model for emotional inference.Nature. 2025 Jul;643(8073):1044-1056. doi: 10.1038/s41586-025-09001-2. Epub 2025 May 14. Nature. 2025. PMID: 40369081
-
Population imaging of internal state circuits relevant to psychiatric disease: a review.Neurophotonics. 2025 Jan;12(Suppl 1):S14607. doi: 10.1117/1.NPh.12.S1.S14607. Epub 2025 Jan 28. Neurophotonics. 2025. PMID: 39872404 Free PMC article. Review.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
