

This is a preprint.
A comprehensive study of SARS-CoV-2 main protease (Mpro) inhibitor-resistant mutants selected in a VSV-based system
- PMID: 37808638
- PMCID: PMC10557589
- DOI: 10.1101/2023.09.22.558628
A comprehensive study of SARS-CoV-2 main protease (Mpro) inhibitor-resistant mutants selected in a VSV-based system
Update in
-
A comprehensive study of SARS-CoV-2 main protease (Mpro) inhibitor-resistant mutants selected in a VSV-based system.PLoS Pathog. 2024 Sep 11;20(9):e1012522. doi: 10.1371/journal.ppat.1012522. eCollection 2024 Sep. PLoS Pathog. 2024. PMID: 39259728 Free PMC article.
Abstract
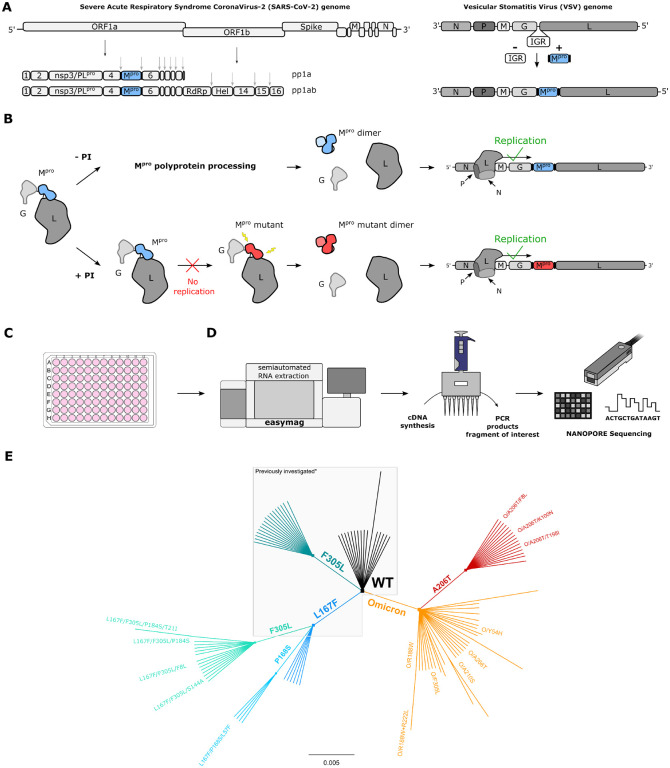
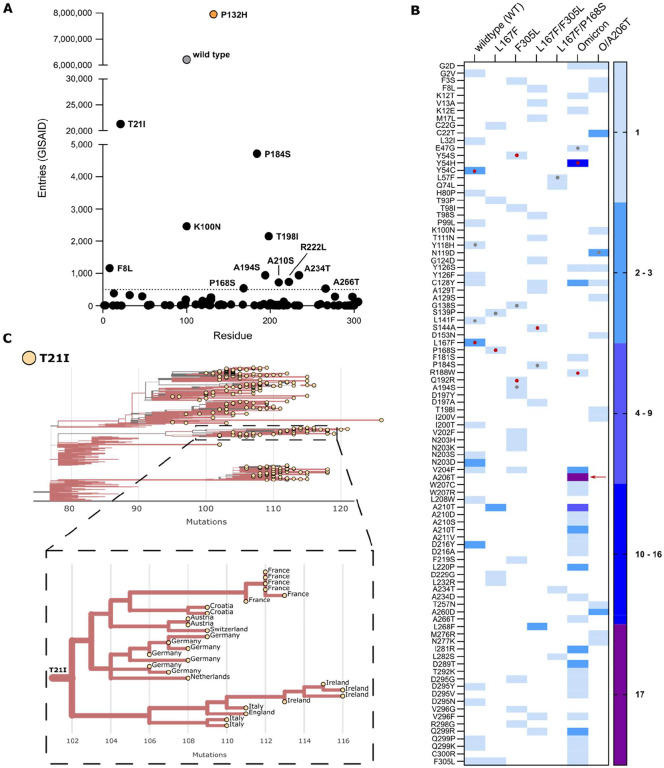
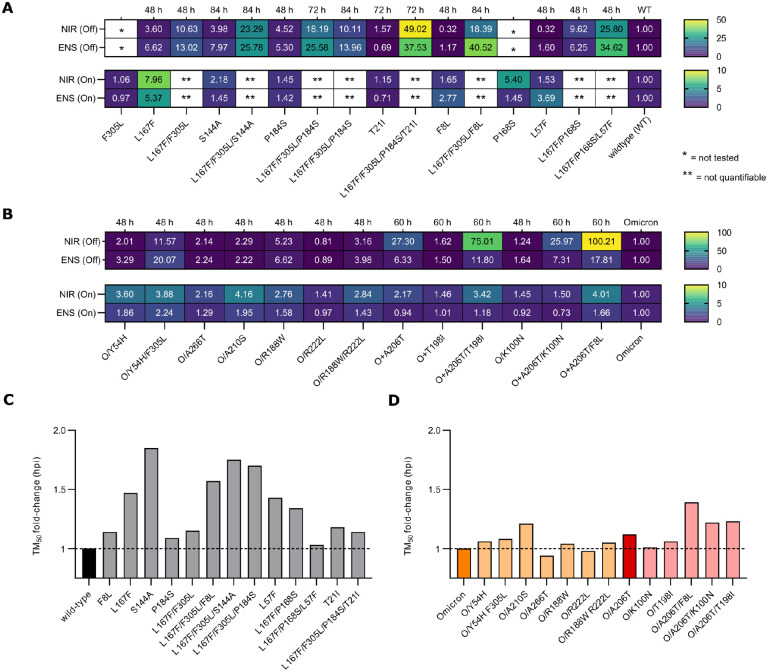
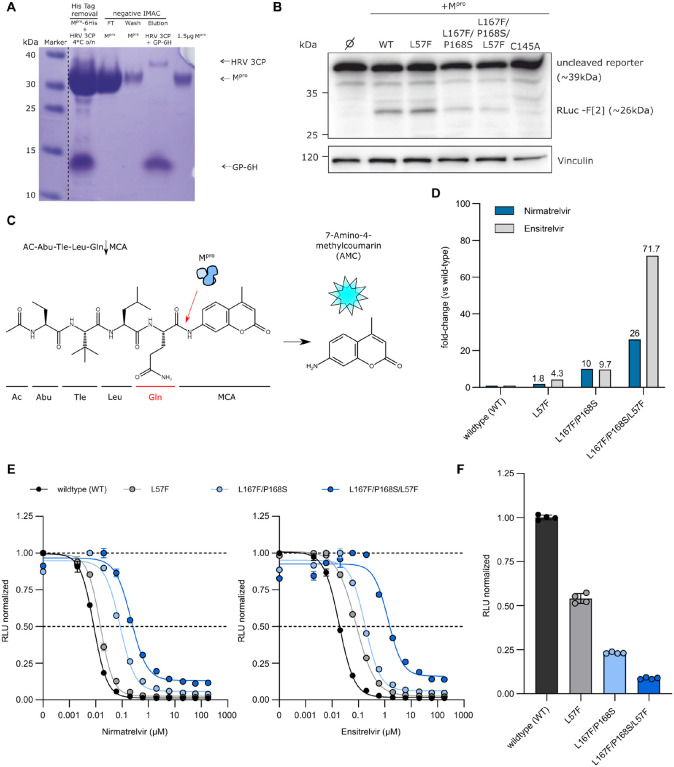
Nirmatrelvir was the first protease inhibitor (PI) specifically developed against the SARS-CoV-2 main protease (3CLpro/Mpro) and licensed for clinical use. As SARS-CoV-2 continues to spread, variants resistant to nirmatrelvir and other currently available treatments are likely to arise. This study aimed to identify and characterize mutations that confer resistance to nirmatrelvir. To safely generate Mpro resistance mutations, we passaged a previously developed, chimeric vesicular stomatitis virus (VSV-Mpro) with increasing, yet suboptimal concentrations of nirmatrelvir. Using Wuhan-1 and Omicron Mpro variants, we selected a large set of mutants. Some mutations are frequently present in GISAID, suggesting their relevance in SARS-CoV-2. The resistance phenotype of a subset of mutations was characterized against clinically available PIs (nirmatrelvir and ensitrelvir) with cell-based and biochemical assays. Moreover, we showed the putative molecular mechanism of resistance based on in silico molecular modelling. These findings have implications on the development of future generation Mpro inhibitors, will help to understand SARS-CoV-2 protease-inhibitor-resistance mechanisms and show the relevance of specific mutations in the clinic, thereby informing treatment decisions.
Keywords: Antimicrobial resistance; Omicron; SARS-CoV-2; ensitrelvir; nirmatrelvir; non-gain-of-function research; protease inhibitor resistance mutations; selection pressure experiments.
Conflict of interest statement
D.v.L. is founder of ViraTherapeutics GmbH. D.v.L serves as a scientific advisor to Boehringer Ingelheim and Pharma KG. E.H. and D.v.L have received an Austrian Science Fund (FWF) grant in the special call “SARS urgent funding”. E.H. is a registered consultant at Guidepoint. D. Bante holds stocks of Pfizer Inc. and Oxford Nanopore Technologies plc. S.A.M and R.S.H. are inventors on the patent application “Live cell assay for protease inhibition”, application number WO/2022/094463. All other authors declare they have no competing interests.
Figures
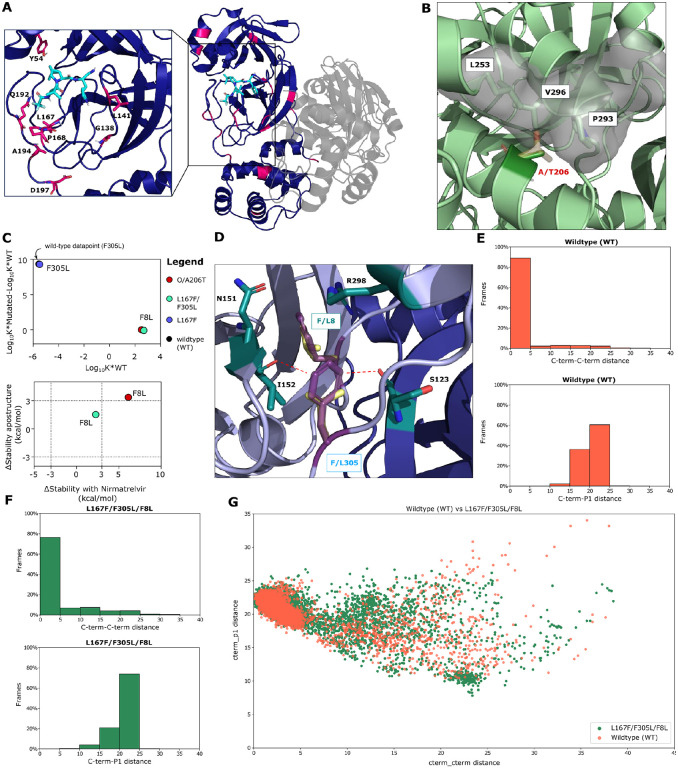
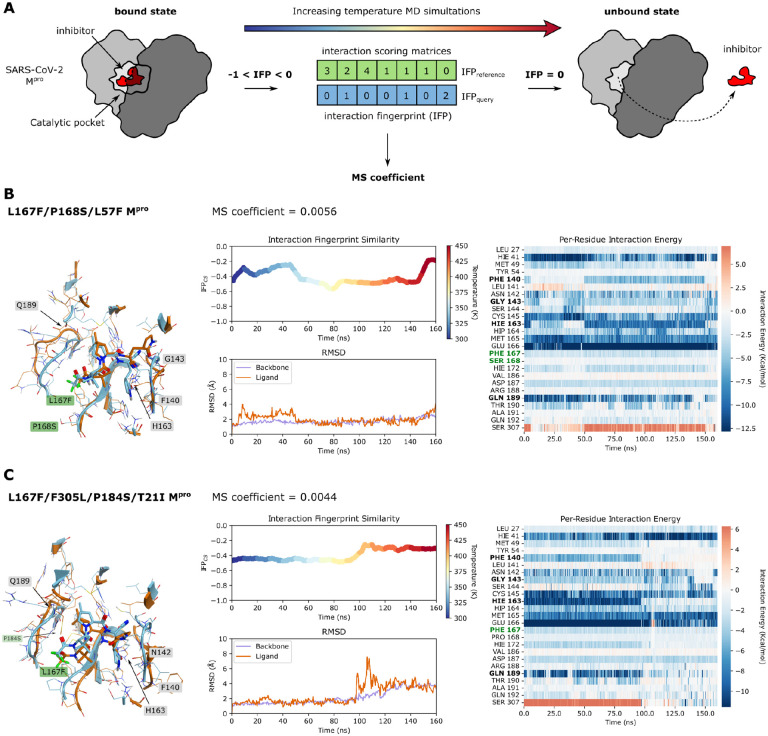
References
-
- Mohammed I., Nauman A., Paul P., Ganesan S., Chen K. H., Jalil S. M. S., Jaouni S. H., Kawas H., Khan W. A., Vattoth A. L., Al-Hashimi Y. A., Fares A., Zeghlache R., Zakaria D., The efficacy and effectiveness of the COVID-19 vaccines in reducing infection, severity, hospitalization, and mortality: a systematic review. Hum. Vaccines Immunother. 18 (2022), doi:10.1080/21645515.2022.2027160. - DOI - PMC - PubMed
-
- Davies N. G., Abbott S., Barnard R. C., Jarvis C. I., Kucharski A. J., Munday J. D., Pearson C. A. B., Russell T. W., Tully D. C., Washburne A. D., Wenseleers T., Gimma A., Waites W., Wong K. L. M., van Zandvoort K., Silverman J. D., Diaz-Ordaz K., Keogh R., Eggo R. M., Funk S., Jit M., Atkins K. E., Edmunds W. J., Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England. Science (80-. ). 372 (2021), doi:10.1126/science.abg3055. - DOI - PMC - PubMed
-
- Kustin T., Harel N., Finkel U., Perchik S., Harari S., Tahor M., Caspi I., Levy R., Leshchinsky M., Ken Dror S., Bergerzon G., Gadban H., Gadban F., Eliassian E., Shimron O., Saleh L., Ben-Zvi H., Keren Taraday E., Amichay D., Ben-Dor A., Sagas D., Strauss M., Shemer Avni Y., Huppert A., Kepten E., Balicer R. D., Netzer D., Ben-Shachar S., Stern A., Evidence for increased breakthrough rates of SARS-CoV-2 variants of concern in BNT162b2-mRNA-vaccinated individuals. Nat. Med. 27, 1379–1384 (2021). - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous