

This is a preprint.
Nuclear lamin A-associated proteins are required for centromere assembly
- PMID: 37808683
- PMCID: PMC10557622
- DOI: 10.1101/2023.09.25.559341
Nuclear lamin A-associated proteins are required for centromere assembly
Abstract
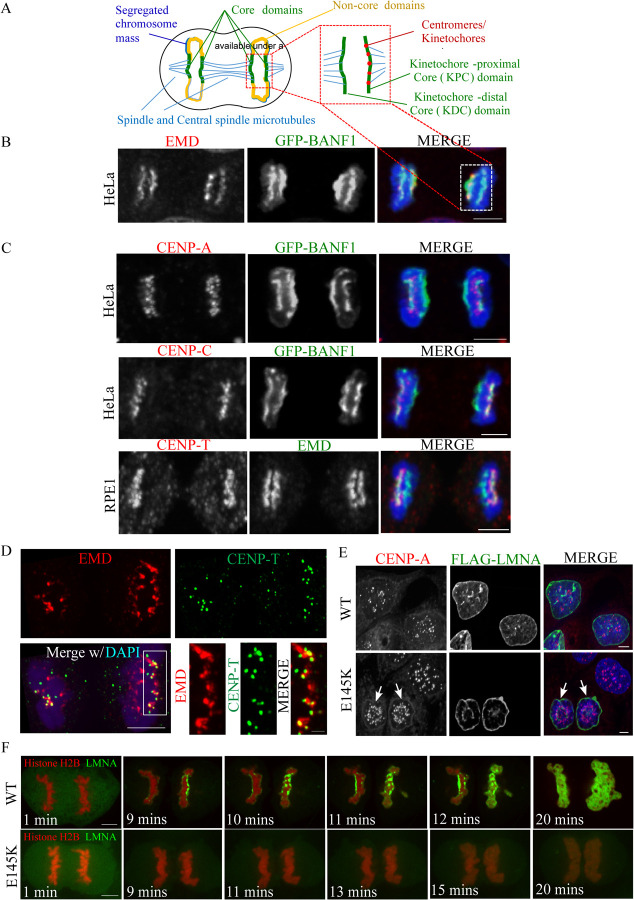
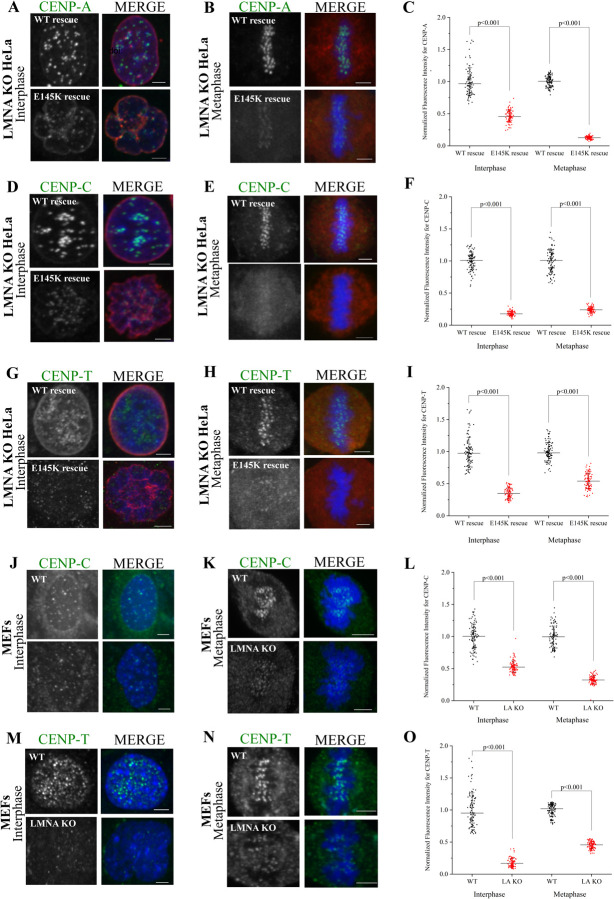
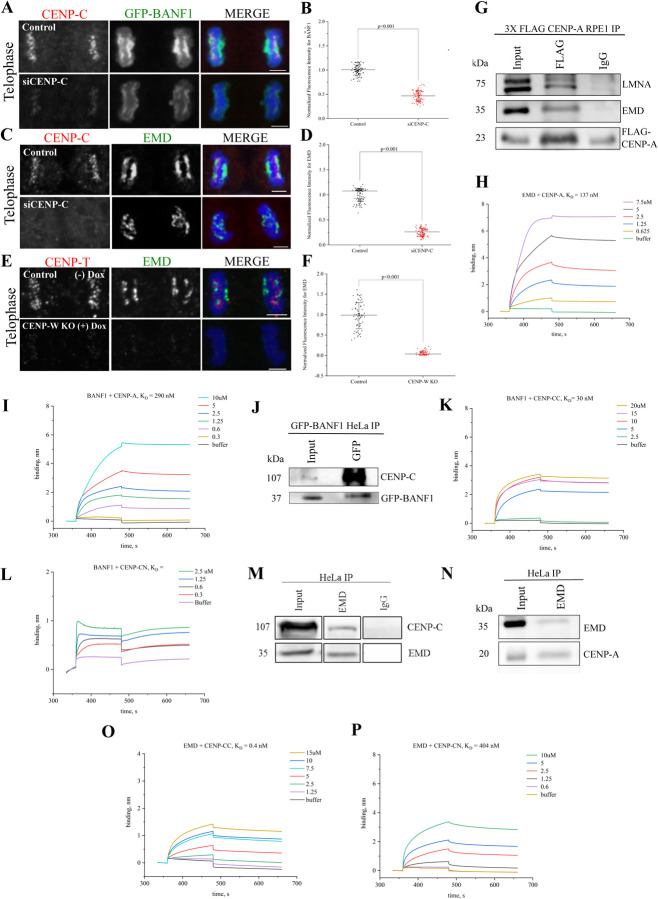
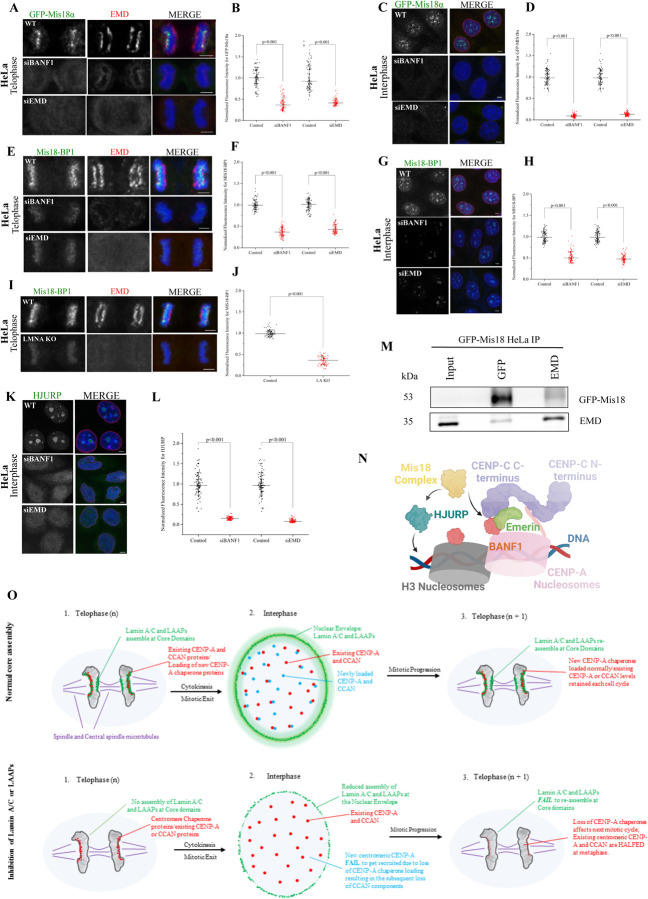
Many Lamin A-associated proteins (LAAP's) that are key constituents of the nuclear envelope (NE), assemble at the "core" domains of chromosomes during NE reformation and mitotic exit. However, the identity and function of the chromosomal core domains remain ill-defined. Here, we show that a distinct section of the core domain overlaps with the centromeres/kinetochores of chromosomes during mitotic telophase. The core domain can thus be demarcated into a kinetochore proximal core (KPC) on one side of the segregated chromosomes and the kinetochore distal core (KDC) on the opposite side, close to the central spindle. We next tested if centromere assembly is connected to NE re-formation. We find that centromere assembly is markedly perturbed after inhibiting the function of LMNA and the core-localized LAAPs, BANF1 and Emerin. We also find that the LAAPs exhibit multiple biochemical interactions with the centromere and inner kinetochore proteins. Consistent with this, normal mitotic progression and chromosome segregation was severely impeded after inhibiting LAAP function. Intriguingly, the inhibition of centromere function also interferes with the assembly of LAAP components at the core domain, suggesting a mutual dependence of LAAP and centromeres for their assembly at the core domains. Finally, we find that the localization of key proteins involved in the centromeric loading of CENP-A, including the Mis18 complex and HJURP were markedly affected in LAAP-inhibited cells. Our evidence points to a model where LAAP assembly at the core domain serves a key function in loading new copies of centromeric proteins during or immediately after mitotic exit.
Keywords: BANF1; CENP-A; Centromere; Emerin; LaminA; Mitosis.
Conflict of interest statement
Conflict of Interest The authors declare no competing financial interests.
Figures
References
-
- Takeuchi K., and Fukagawa T. (2012). Molecular architecture of vertebrate kinetochores. Experimental cell research 318, 1367–1374. - PubMed
-
- Hori T., and Fukagawa T. (2012). Establishment of the vertebrate kinetochores. Chromosome Res 20, 547–561. - PubMed
-
- Ray-Gallet D., and Almouzni G. (2021). The Histone H3 Family and Its Deposition Pathways. Adv Exp Med Biol 1283, 17–42. - PubMed
-
- Cheeseman I.M., and Desai A. (2008). Molecular architecture of the kinetochore-microtubule interface. Nat Rev Mol Cell Biol 9, 33–46. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous