

This is a preprint.
Defective mesenchymal Bmpr1a-mediated BMP signaling causes congenital pulmonary cysts
- PMID: 37808788
- PMCID: PMC10557633
- DOI: 10.1101/2023.09.26.559527
Defective mesenchymal Bmpr1a-mediated BMP signaling causes congenital pulmonary cysts
Update in
-
Defective mesenchymal Bmpr1a-mediated BMP signaling causes congenital pulmonary cysts.Elife. 2024 Jun 10;12:RP91876. doi: 10.7554/eLife.91876. Elife. 2024. PMID: 38856718 Free PMC article.
Abstract
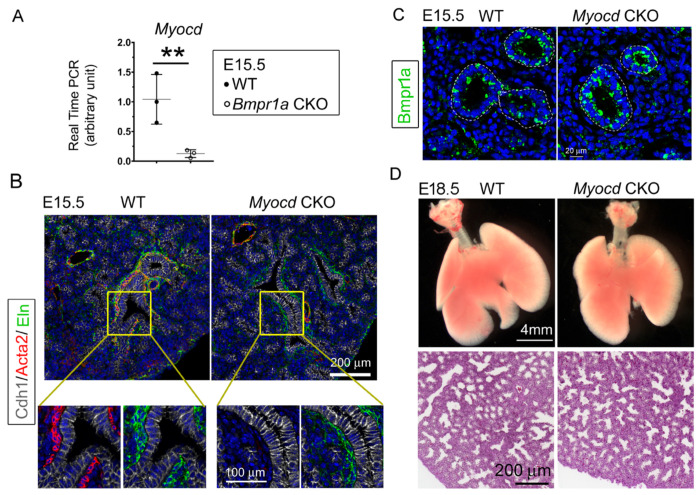
Abnormal lung development can cause congenital pulmonary cysts, the mechanisms of which remain largely unknown. Although the cystic lesions are believed to result directly from disrupted airway epithelial cell growth, the extent to which developmental defects in lung mesenchymal cells contribute to abnormal airway epithelial cell growth and subsequent cystic lesions has not been thoroughly examined. In the present study, we dissected the roles of BMP receptor 1a (Bmpr1a)-mediated BMP signaling in lung mesenchyme during prenatal lung development and discovered that abrogation of mesenchymal Bmpr1a disrupted normal lung branching morphogenesis, leading to the formation of prenatal pulmonary cystic lesions. Severe deficiency of airway smooth muscle cells and subepithelial elastin fibers were found in the cystic airways of the mesenchymal Bmpr1a knockout lungs. In addition, ectopic mesenchymal expression of BMP ligands and airway epithelial perturbation of the Sox2-Sox9 proximal-distal axis were detected in the mesenchymal Bmpr1a knockout lungs. However, deletion of Smad1/5, two major BMP signaling downstream effectors, from the lung mesenchyme did not phenocopy the cystic abnormalities observed in the mesenchymal Bmpr1a knockout lungs, suggesting that a Smad-independent mechanism contributes to prenatal pulmonary cystic lesions. These findings reveal for the first time the role of mesenchymal BMP signaling in lung development and a potential pathogenic mechanism underlying congenital pulmonary cysts.
Keywords: Airway smooth muscle cells; BMP signaling; Bmpr1a; Lung branching morphogenesis; Lung development; Lung mesenchymal cells; Pulmonary cysts.
Figures






References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials