

This is a preprint.
CELLULAR RNA INTERACTS WITH MAVS TO PROMOTE ANTIVIRAL SIGNALING
- PMID: 37808873
- PMCID: PMC10557580
- DOI: 10.1101/2023.09.25.559083
CELLULAR RNA INTERACTS WITH MAVS TO PROMOTE ANTIVIRAL SIGNALING
Update in
-
Cellular RNA interacts with MAVS to promote antiviral signaling.Science. 2024 Dec 20;386(6728):eadl0429. doi: 10.1126/science.adl0429. Epub 2024 Dec 20. Science. 2024. PMID: 39700280
Abstract
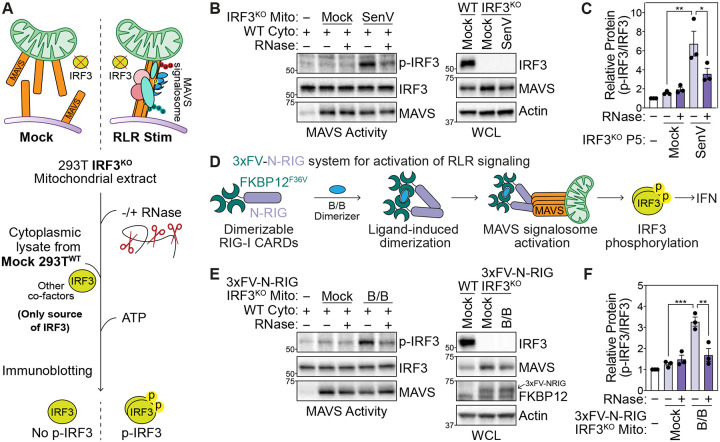
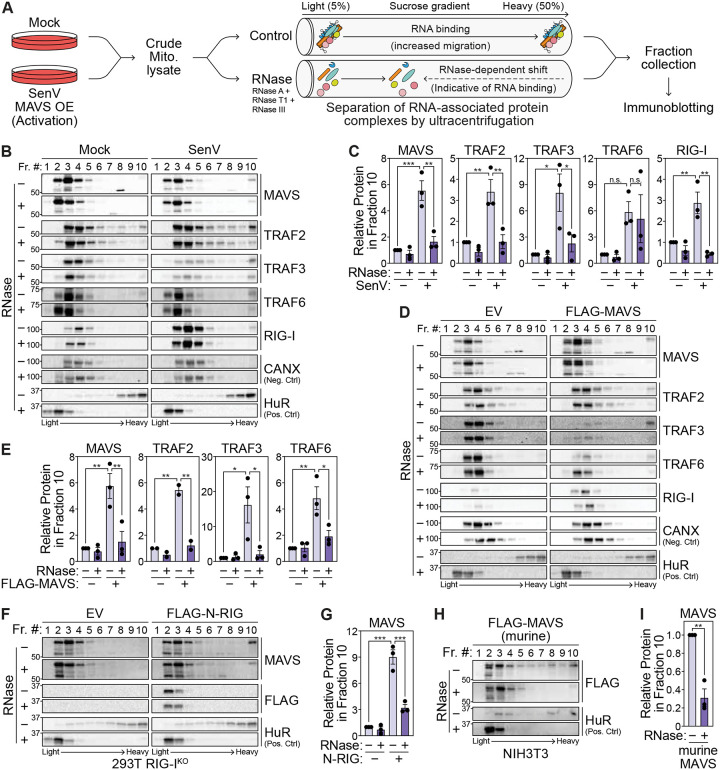
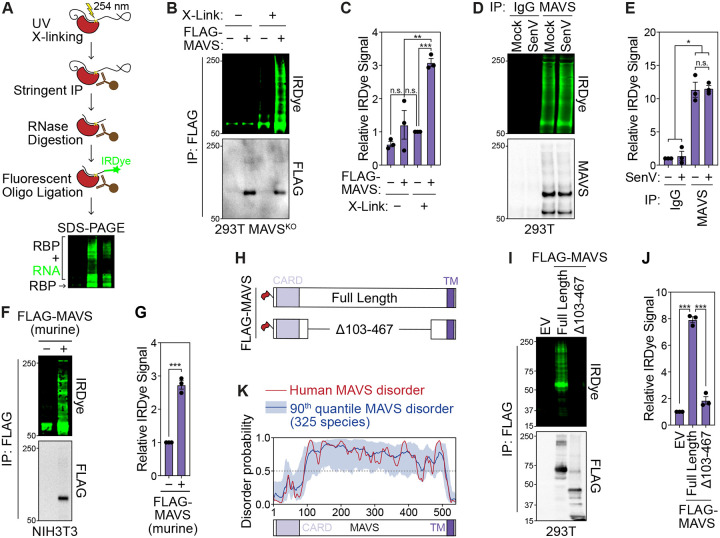
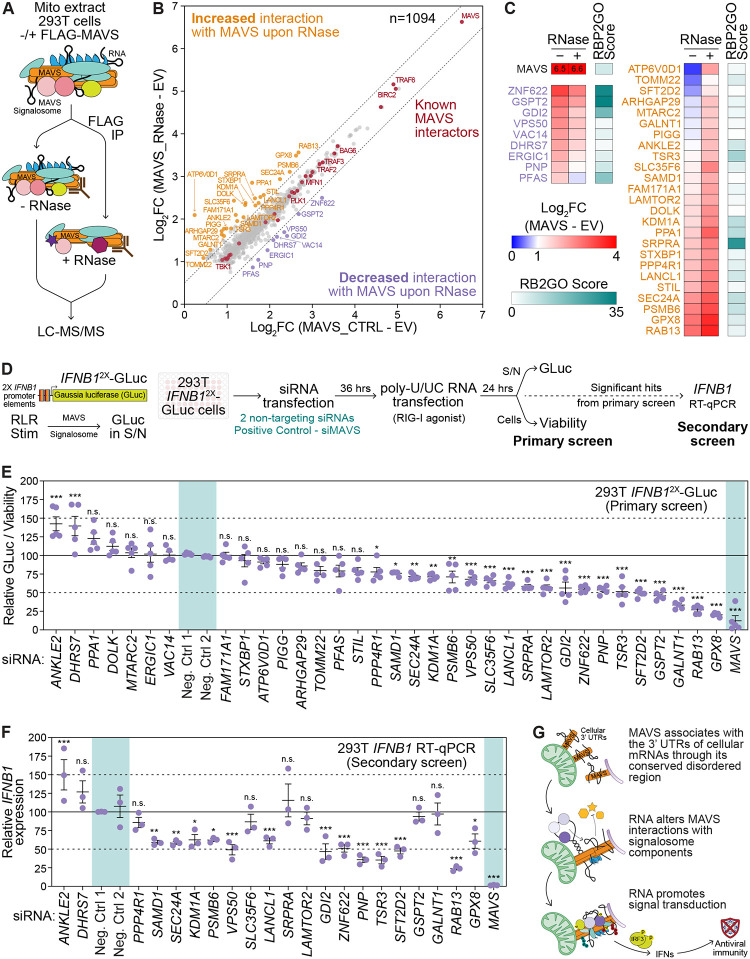
Immune signaling needs to be well-regulated to promote clearance of pathogens, while preventing aberrant inflammation. Interferons (IFNs) and antiviral genes are activated by the detection of viral RNA by RIG-I-like receptors (RLRs). Signal transduction downstream of RLRs proceeds through a multi-protein complex organized around the central adaptor protein MAVS. Recent work has shown that protein complex function can be modulated by RNA molecules providing allosteric regulation or acting as molecular guides or scaffolds. Thus, we hypothesized that RNA plays a role in organizing MAVS signaling platforms. Here, we show that MAVS, through its central intrinsically disordered domain, directly interacts with the 3' untranslated regions of cellular mRNAs. Importantly, elimination of RNA by RNase treatment disrupts the MAVS signalosome, including newly identified regulators of RLR signaling, and inhibits phosphorylation of the transcription factor IRF3. This supports the hypothesis that RNA molecules scaffold proteins in the MAVS signalosome to induce IFNs. Together, this work uncovers a function for cellular RNA in promoting signaling through MAVS and highlights a generalizable principle of RNA regulatory control of cytoplasmic immune signaling complexes.
Conflict of interest statement
COMPETING INTERESTS M.G. is a founder and shareholder in Kineta, Inc., and of HDT Bio.
Figures

References
-
- Kato H., Takeuchi O., Sato S., Yoneyama M., Yamamoto M., Matsui K., Uematsu S., Jung A., Kawai T., Ishii K. J., Yamaguchi O., Otsu K., Tsujimura T., Koh C.-S., Reis e Sousa C., Matsuura Y., Fujita T., Akira S., Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature. 441, 101–105 (2006). - PubMed
Publication types
Grants and funding
- R01 AI153246/AI/NIAID NIH HHS/United States
- R21 AI158788/AI/NIAID NIH HHS/United States
- R01 GM129090/GM/NIGMS NIH HHS/United States
- T32 AI106677/AI/NIAID NIH HHS/United States
- P51 OD010425/OD/NIH HHS/United States
- R01 AI155512/AI/NIAID NIH HHS/United States
- R01 AI175724/AI/NIAID NIH HHS/United States
- R21 AI141823/AI/NIAID NIH HHS/United States
- R01 AI145296/AI/NIAID NIH HHS/United States
- R21 AI176442/AI/NIAID NIH HHS/United States
- S10 OD021502/OD/NIH HHS/United States
- K99 AI175483/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous