

Breast cancer stem cells tolerate chromosomal instability during tumor progression via c-Jun/AXL stress signaling
- PMID: 37809421
- PMCID: PMC10559946
- DOI: 10.1016/j.heliyon.2023.e20182
Breast cancer stem cells tolerate chromosomal instability during tumor progression via c-Jun/AXL stress signaling
Abstract
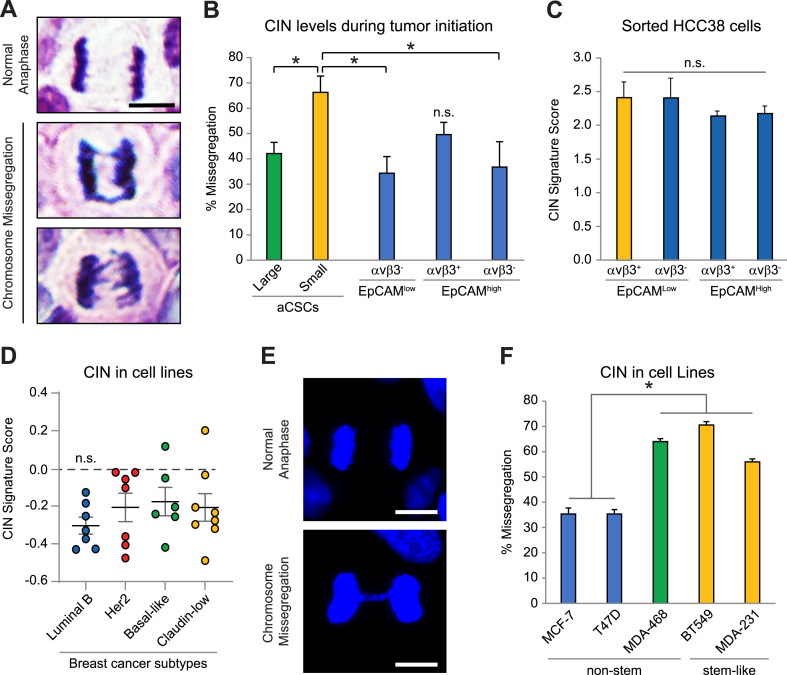
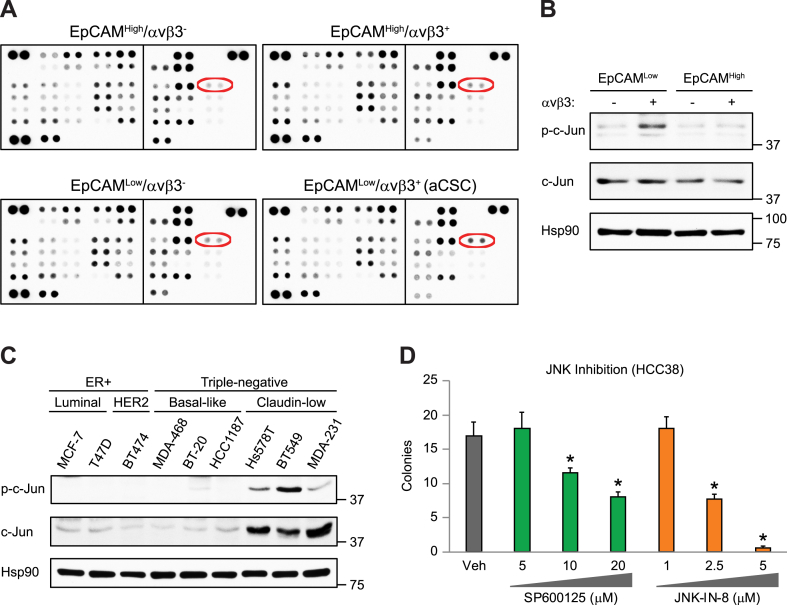
Chromosomal instability (CIN) is critical for tumor evolution, yet its relationship with stemness is unclear. Here, we describe CIN as a key stress induced during tumor initiation that is uniquely tolerated by breast cancer stem cells in an activated signaling state (aCSCs). While we noted elevated CIN specifically in tumors from aCSCs, this was not intrinsic to these cells, as baseline levels were similar to non-stem cell types. This suggests that CIN is induced during tumor initiation, and that aCSCs can better tolerate this stress. Further, this increased CIN may be transient, as it was only in low-burden aCSC tumors, with levels diminishing in more established disease. Phospho-array profiling revealed specific activation of c-Jun stress signaling in aCSCs, which we hypothesized could induce genes responsible for CIN tolerance. Indeed, we identified AXL as a c-Jun dependent gene enriched in aCSCs that enhances resistance to this stress. Thus, CIN tolerance mediated by c-Jun/AXL signaling may be a defining feature of stemness, contributing to breast cancer progression.
Keywords: Breast cancer; Chromosomal missegregation; Stem cell activation; Stress tolerance; Tumor initiation.
© 2023 The Authors.
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures


References
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
