

Regulation of CTCF loop formation during pancreatic cell differentiation
- PMID: 37813869
- PMCID: PMC10562423
- DOI: 10.1038/s41467-023-41964-6
Regulation of CTCF loop formation during pancreatic cell differentiation
Abstract
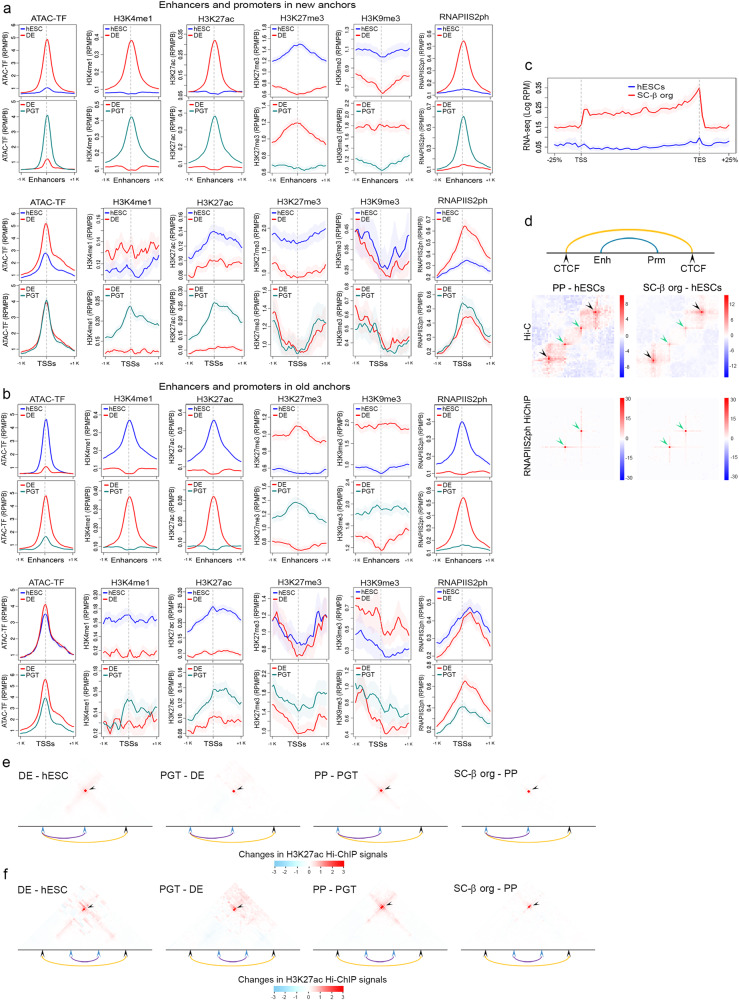
Transcription reprogramming during cell differentiation involves targeting enhancers to genes responsible for establishment of cell fates. To understand the contribution of CTCF-mediated chromatin organization to cell lineage commitment, we analyzed 3D chromatin architecture during the differentiation of human embryonic stem cells into pancreatic islet organoids. We find that CTCF loops are formed and disassembled at different stages of the differentiation process by either recruitment of CTCF to new anchor sites or use of pre-existing sites not previously involved in loop formation. Recruitment of CTCF to new sites in the genome involves demethylation of H3K9me3 to H3K9me2, demethylation of DNA, recruitment of pioneer factors, and positioning of nucleosomes flanking the new CTCF sites. Existing CTCF sites not involved in loop formation become functional loop anchors via the establishment of new cohesin loading sites containing NIPBL and YY1 at sites between the new anchors. In both cases, formation of new CTCF loops leads to strengthening of enhancer promoter interactions and increased transcription of genes adjacent to loop anchors. These results suggest an important role for CTCF and cohesin in controlling gene expression during cell differentiation.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
