

The accumulation and growth of Pseudomonas aeruginosa on surfaces is modulated by surface mechanics via cyclic-di-GMP signaling
- PMID: 37816780
- PMCID: PMC10564899
- DOI: 10.1038/s41522-023-00436-x
The accumulation and growth of Pseudomonas aeruginosa on surfaces is modulated by surface mechanics via cyclic-di-GMP signaling
Abstract
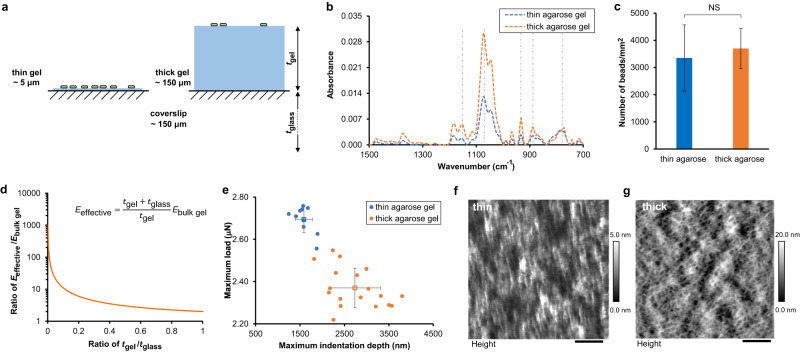
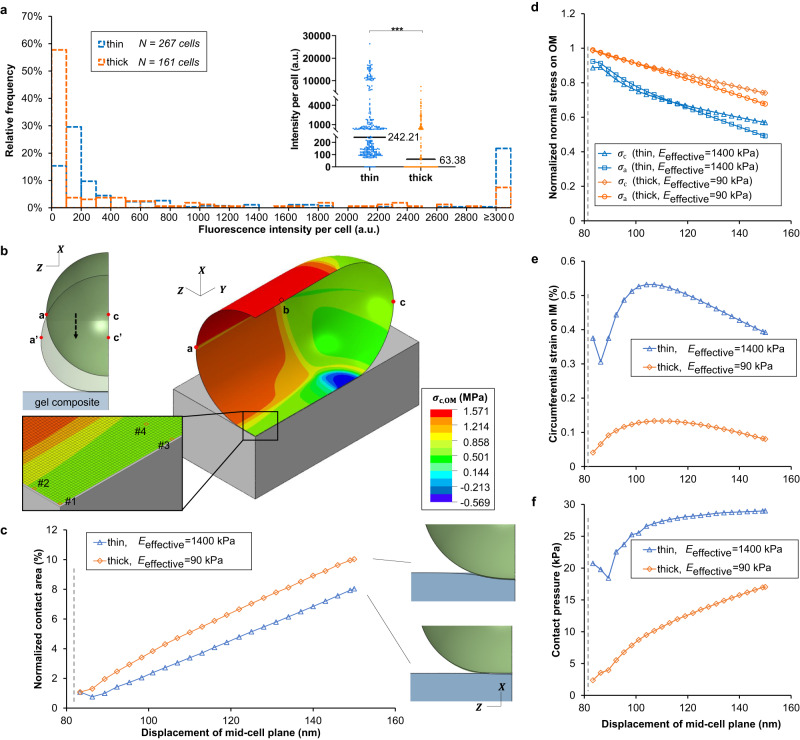
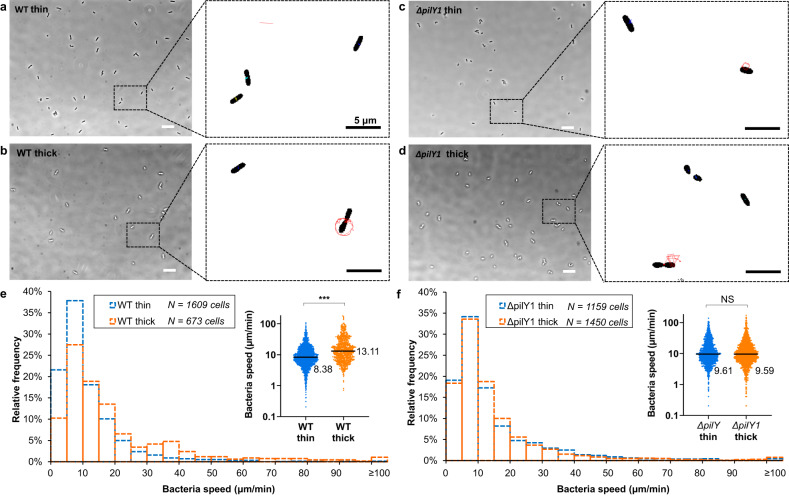
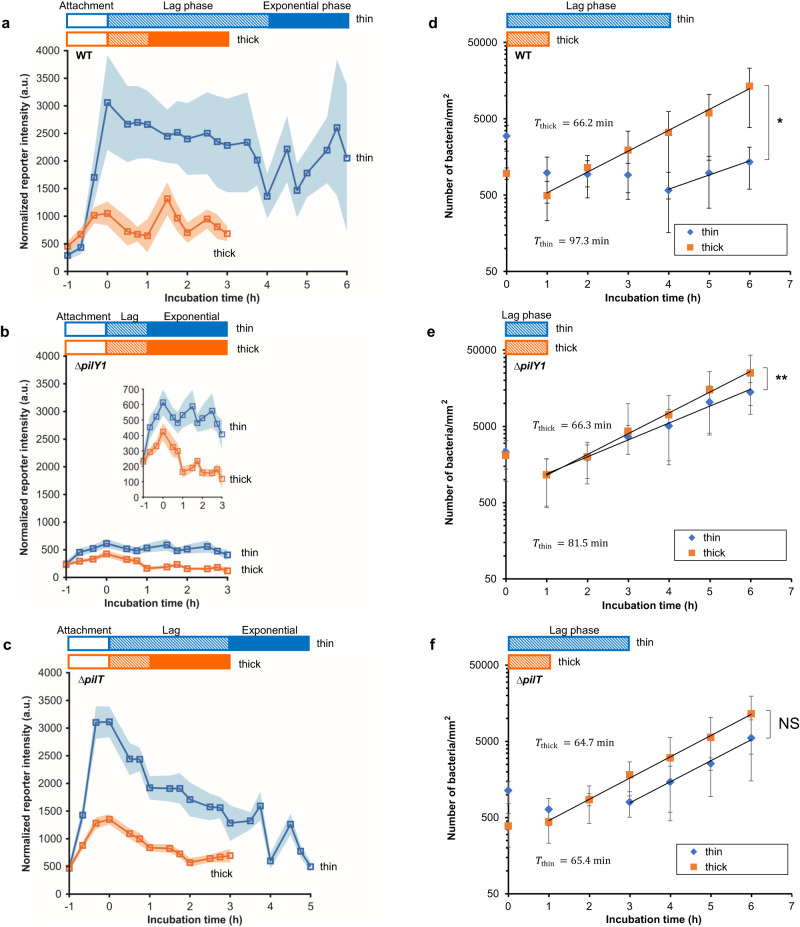
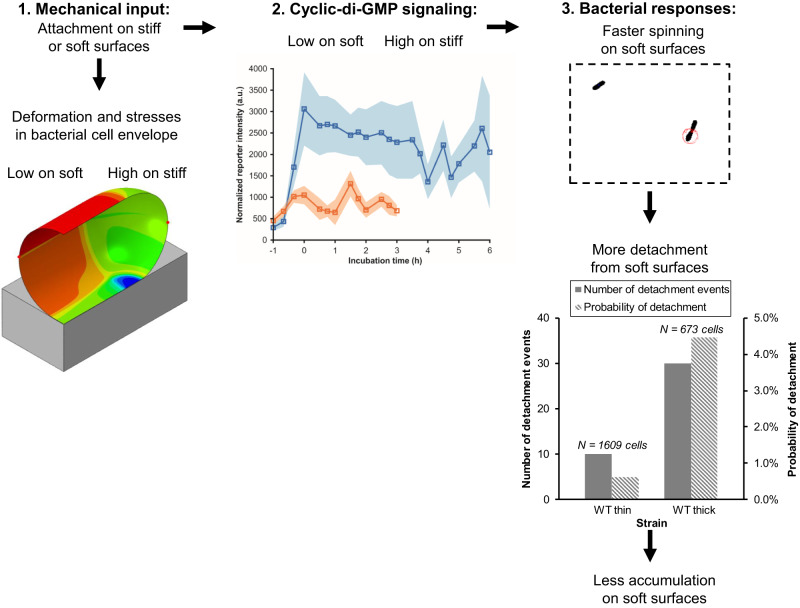
Attachment of bacteria onto a surface, consequent signaling, and accumulation and growth of the surface-bound bacterial population are key initial steps in the formation of pathogenic biofilms. While recent reports have hinted that surface mechanics may affect the accumulation of bacteria on that surface, the processes that underlie bacterial perception of surface mechanics and modulation of accumulation in response to surface mechanics remain largely unknown. We use thin and thick hydrogels coated on glass to create composite materials with different mechanics (higher elasticity for thin composites; lower elasticity for thick composites) but with the same surface adhesivity and chemistry. The mechanical cue stemming from surface mechanics is elucidated using experiments with the opportunistic human pathogen Pseudomonas aeruginosa combined with finite-element modeling. Adhesion to thin composites results in greater changes in mechanical stress and strain in the bacterial envelope than does adhesion to thick composites with identical surface chemistry. Using quantitative microscopy, we find that adhesion to thin composites also results in higher cyclic-di-GMP levels, which in turn result in lower motility and less detachment, and thus greater accumulation of bacteria on the surface than does adhesion to thick composites. Mechanics-dependent c-di-GMP production is mediated by the cell-surface-exposed protein PilY1. The biofilm lag phase, which is longer for bacterial populations on thin composites than on thick composites, is also mediated by PilY1. This study shows clear evidence that bacteria actively regulate differential accumulation on surfaces of different stiffnesses via perceiving varied mechanical stress and strain upon surface engagement.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures

Update of
-
Bacterial mechanosensing of surface stiffness promotes signaling and growth leading to biofilm formation by Pseudomonas aeruginosa.bioRxiv [Preprint]. 2023 Jan 27:2023.01.26.525810. doi: 10.1101/2023.01.26.525810. bioRxiv. 2023. Update in: NPJ Biofilms Microbiomes. 2023 Oct 10;9(1):78. doi: 10.1038/s41522-023-00436-x. PMID: 36747833 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
