

STING activation in TET2-mutated hematopoietic stem/progenitor cells contributes to the increased self-renewal and neoplastic transformation
- PMID: 37816954
- PMCID: PMC10681905
- DOI: 10.1038/s41375-023-02055-z
STING activation in TET2-mutated hematopoietic stem/progenitor cells contributes to the increased self-renewal and neoplastic transformation
Abstract
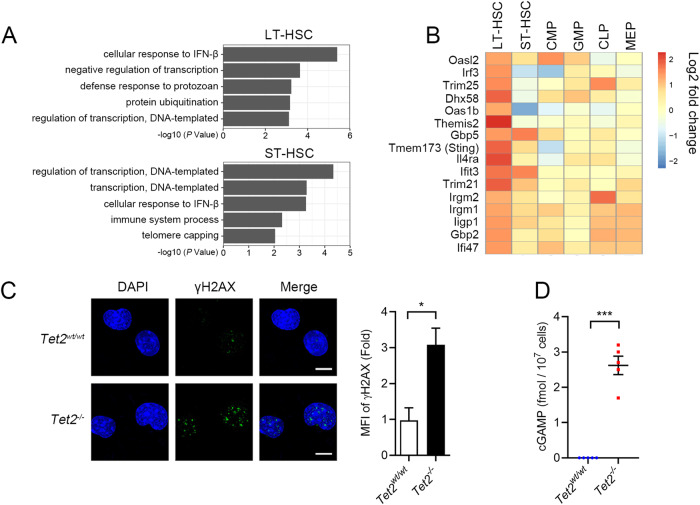
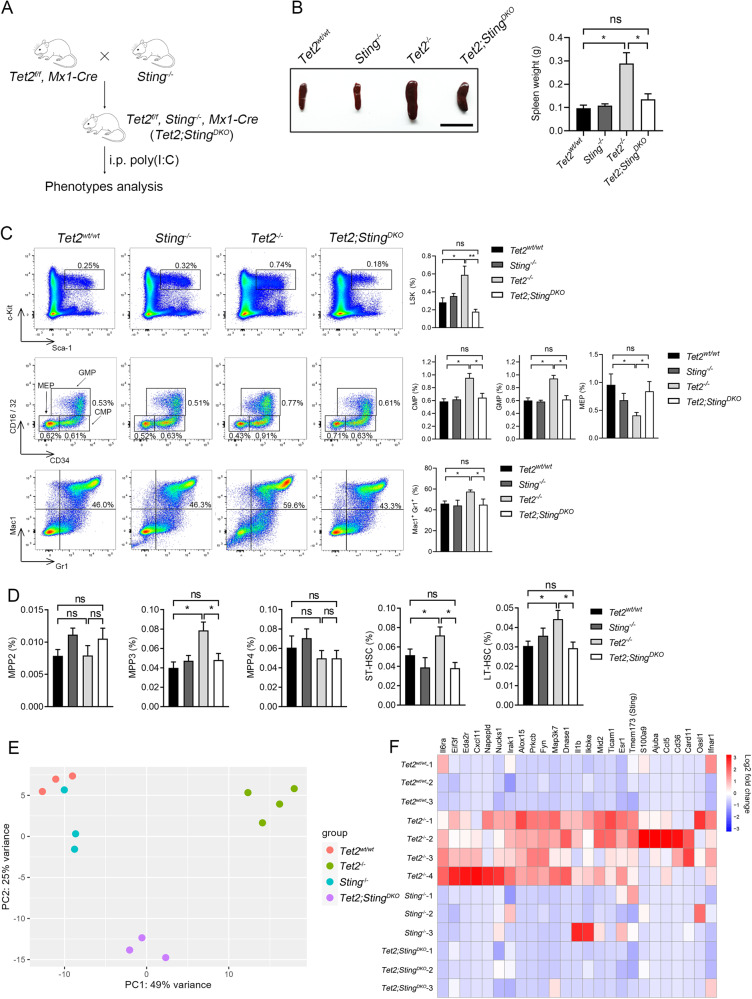
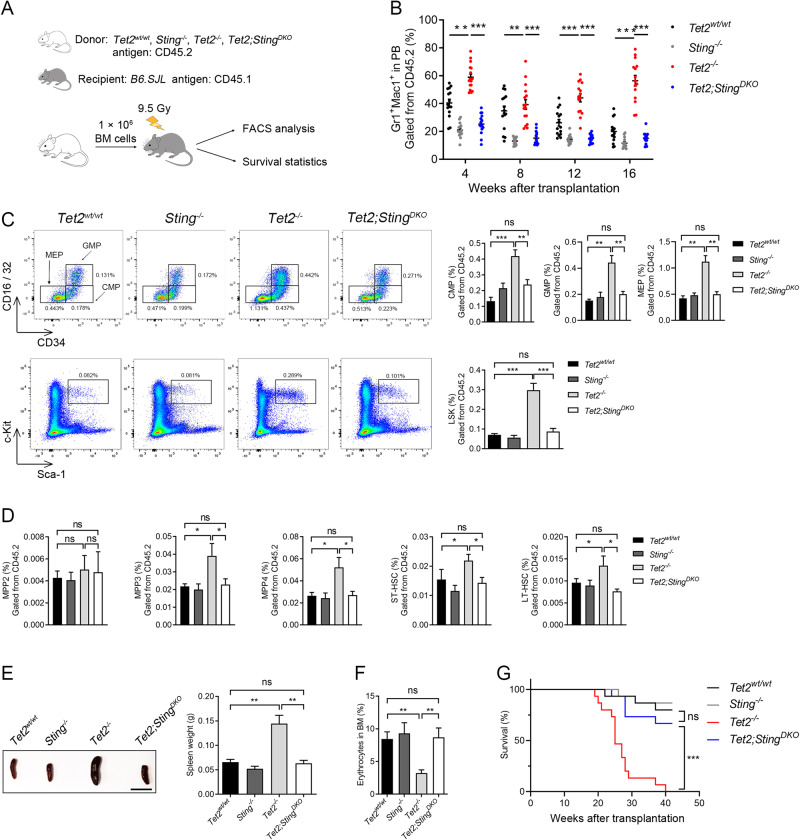
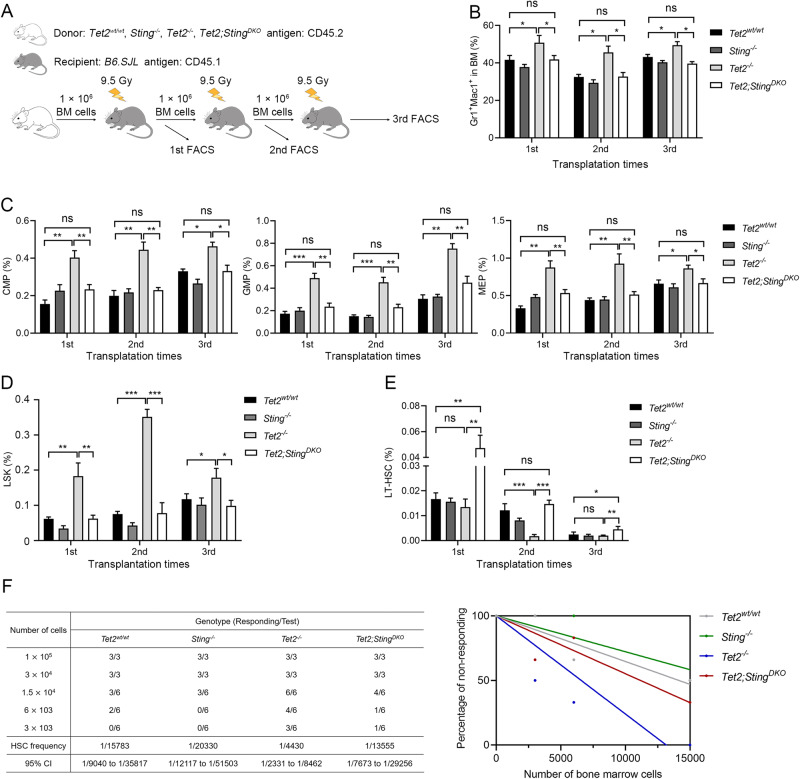
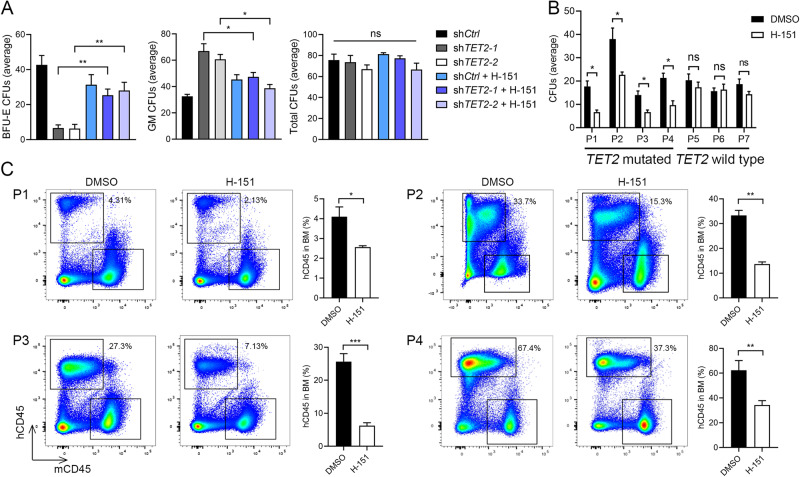
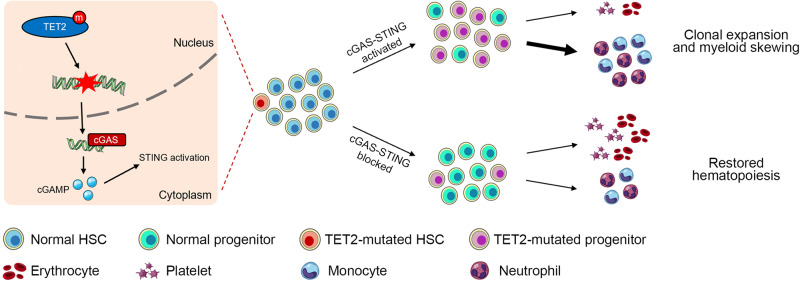
Somatic loss-of-function mutations of the dioxygenase Ten-eleven translocation-2 (TET2) occur frequently in individuals with clonal hematopoiesis (CH) and acute myeloid leukemia (AML). These common hematopoietic disorders can be recapitulated in mouse models. However, the underlying mechanisms by which the deficiency in TET2 promotes these disorders remain unclear. Here we show that the cyclic guanosine monophosphate-adenosine monophosphate synthase (cGAS)-stimulator of interferon genes (STING) pathway is activated to mediate the effect of TET2 deficiency in dysregulated hematopoiesis in mouse models. DNA damage arising in Tet2-deficient hematopoietic stem/progenitor cells (HSPCs) leads to activation of the cGAS-STING pathway which in turn promotes the enhanced self-renewal and development of CH. Notably, both pharmacological inhibition and genetic deletion of STING suppresses Tet2 mutation-induced aberrant hematopoiesis. In patient-derived xenograft (PDX) models, STING inhibition specifically attenuates the proliferation of leukemia cells from TET2-mutated individuals. These observations suggest that the development of CH associated with TET2 mutations is powered through chronic inflammation dependent on the activated cGAS-STING pathway and that STING may represent a potential target for intervention of relevant hematopoietic diseases.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
