

The landscape of human SVA retrotransposons
- PMID: 37823611
- PMCID: PMC10681720
- DOI: 10.1093/nar/gkad821
The landscape of human SVA retrotransposons
Abstract
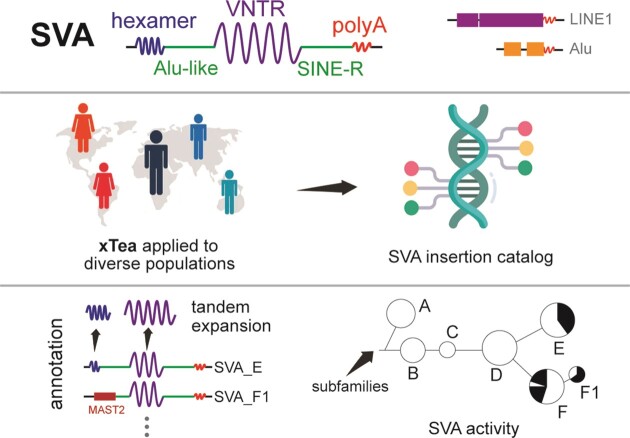
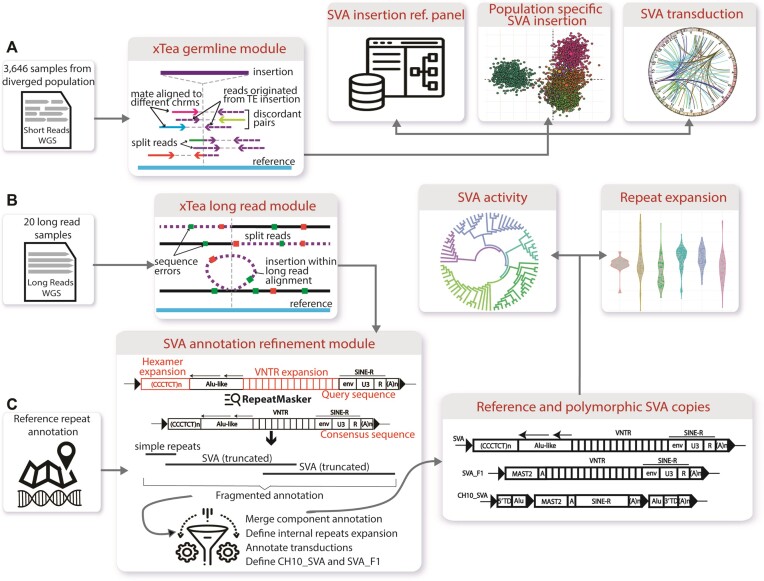
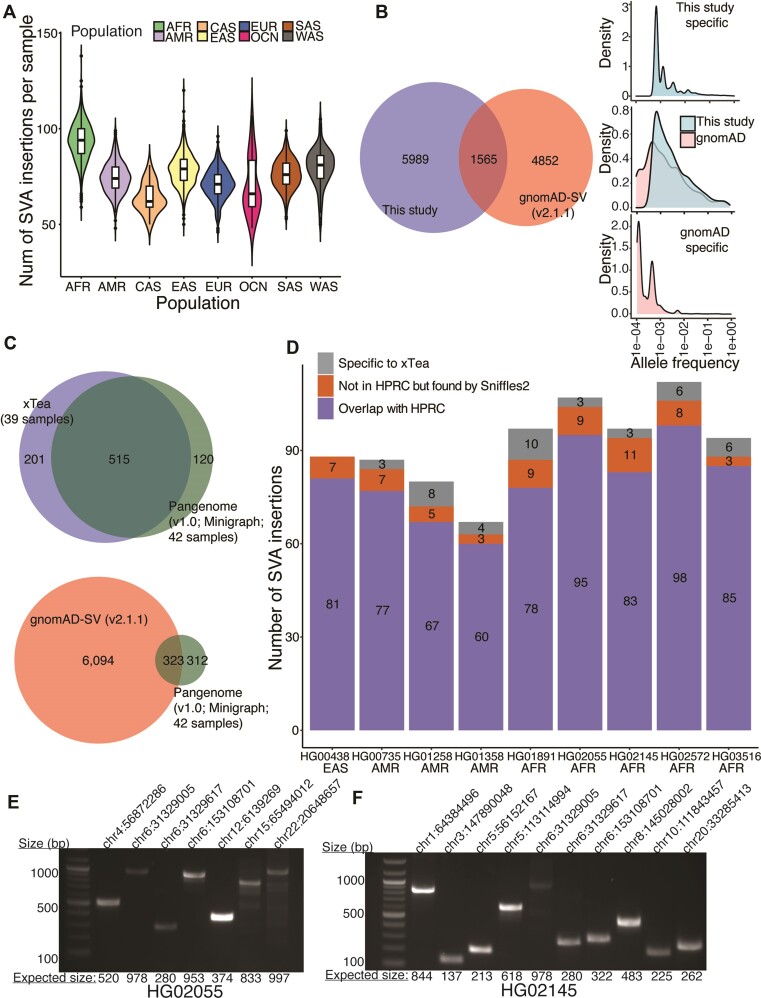
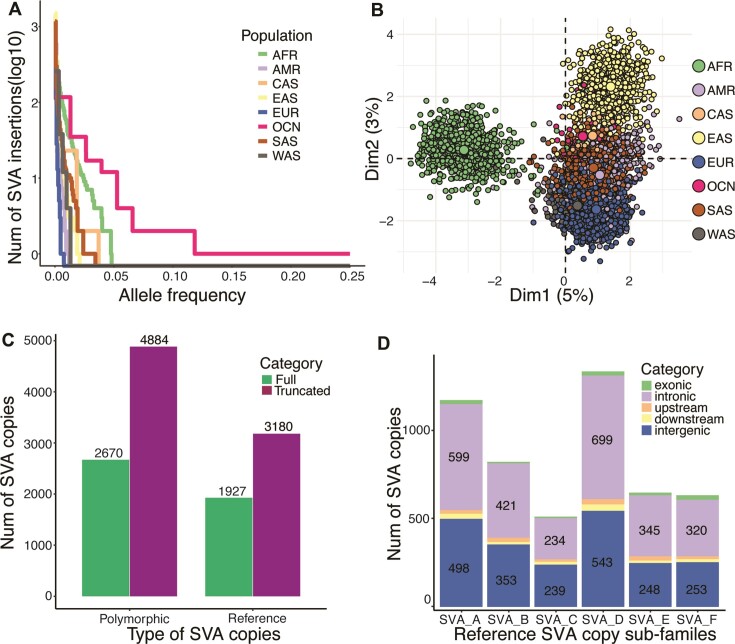
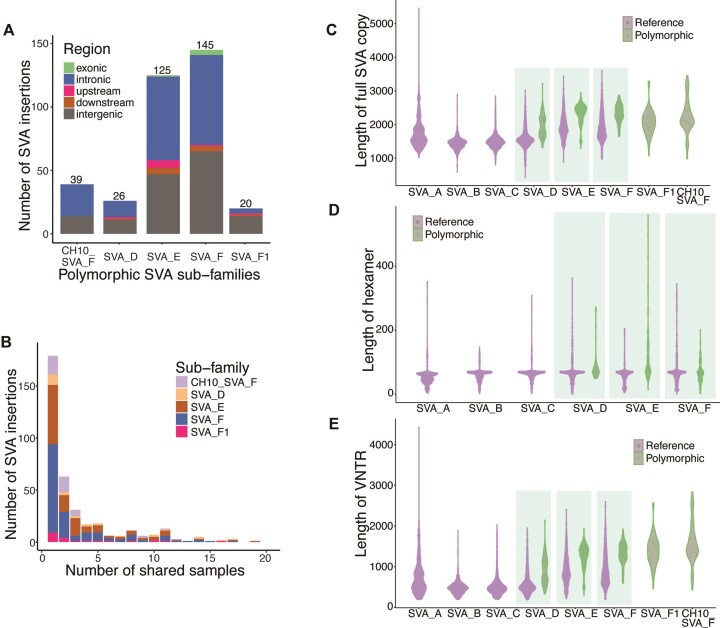
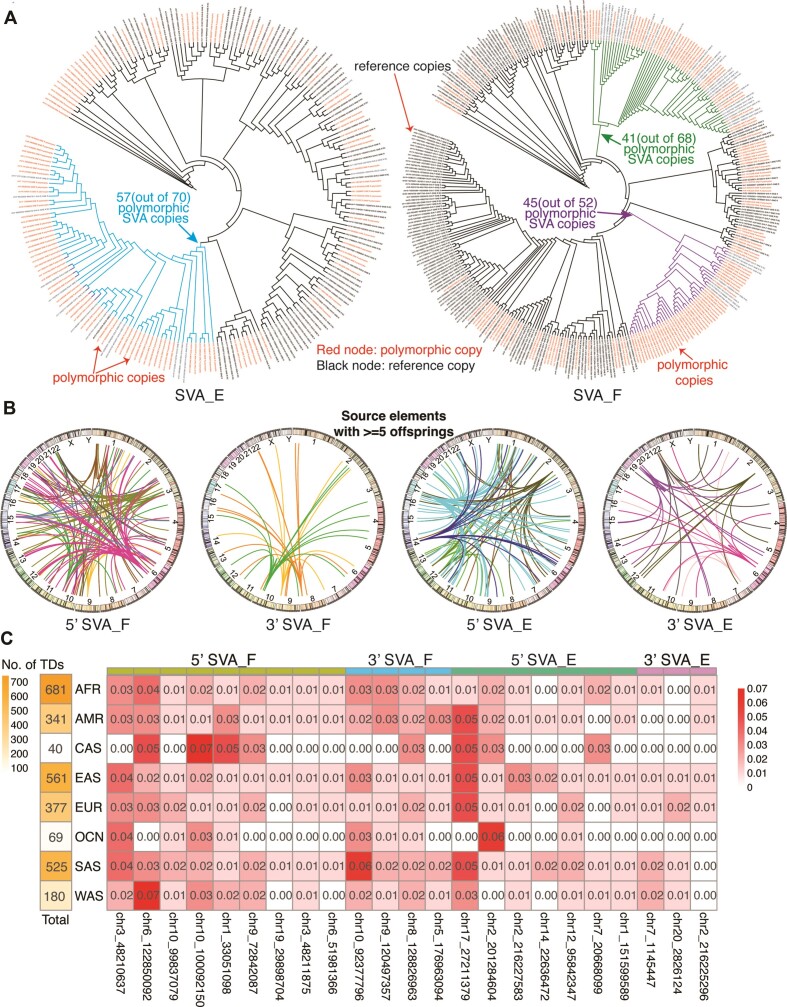
SINE-VNTR-Alu (SVA) retrotransposons are evolutionarily young and still-active transposable elements (TEs) in the human genome. Several pathogenic SVA insertions have been identified that directly mutate host genes to cause neurodegenerative and other types of diseases. However, due to their sequence heterogeneity and complex structures as well as limitations in sequencing techniques and analysis, SVA insertions have been less well studied compared to other mobile element insertions. Here, we identified polymorphic SVA insertions from 3646 whole-genome sequencing (WGS) samples of >150 diverse populations and constructed a polymorphic SVA insertion reference catalog. Using 20 long-read samples, we also assembled reference and polymorphic SVA sequences and characterized the internal hexamer/variable-number-tandem-repeat (VNTR) expansions as well as differing SVA activity for SVA subfamilies and human populations. In addition, we developed a module to annotate both reference and polymorphic SVA copies. By characterizing the landscape of both reference and polymorphic SVA retrotransposons, our study enables more accurate genotyping of these elements and facilitate the discovery of pathogenic SVA insertions.
© The Author(s) 2023. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
A hexamer tandem repeat RNA embedded within an SVA retrotransposon drives R-loop formation and neurodegeneration.Cell Rep. 2025 Jul 22;44(7):115812. doi: 10.1016/j.celrep.2025.115812. Epub 2025 Jun 19. Cell Rep. 2025. PMID: 40540399
-
Polygenic burden of short tandem repeat expansions promotes risk for Alzheimer's disease.Nat Commun. 2025 Jan 28;16(1):1126. doi: 10.1038/s41467-025-56400-0. Nat Commun. 2025. PMID: 39875385 Free PMC article.
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
SVA retrotransposons: Evolution and genetic instability.Semin Cancer Biol. 2010 Aug;20(4):234-45. doi: 10.1016/j.semcancer.2010.04.001. Epub 2010 Apr 21. Semin Cancer Biol. 2010. PMID: 20416380 Free PMC article. Review.
-
Carbon dioxide detection for diagnosis of inadvertent respiratory tract placement of enterogastric tubes in children.Cochrane Database Syst Rev. 2025 Feb 19;2(2):CD011196. doi: 10.1002/14651858.CD011196.pub2. Cochrane Database Syst Rev. 2025. PMID: 39968844
Cited by
-
TSPAN4+ fibroblasts coordinate metastatic niche assembly through migrasome-driven metabolic reprogramming and stromal-immune crosstalk in pancreatic adenocarcinoma.Front Immunol. 2025 May 15;16:1594879. doi: 10.3389/fimmu.2025.1594879. eCollection 2025. Front Immunol. 2025. PMID: 40443671 Free PMC article.
-
Regulation and function of transposable elements in cancer genomes.Cell Mol Life Sci. 2024 Mar 31;81(1):157. doi: 10.1007/s00018-024-05195-2. Cell Mol Life Sci. 2024. PMID: 38556602 Free PMC article. Review.
-
Variable patterns of retrotransposition in different HeLa strains provide mechanistic insights into SINE RNA mobilization processes.Nucleic Acids Res. 2024 Jul 22;52(13):7761-7779. doi: 10.1093/nar/gkae448. Nucleic Acids Res. 2024. PMID: 38850156 Free PMC article.
-
Advanced analysis of retrotransposon variation in the human genome with nanopore sequencing using RetroInspector.Sci Rep. 2025 Apr 25;15(1):14489. doi: 10.1038/s41598-025-98847-7. Sci Rep. 2025. PMID: 40281075 Free PMC article.
-
The landscape of non-reference SINE-VNTR-Alus in amyotrophic lateral sclerosis.Exp Biol Med (Maywood). 2025 May 29;250:10600. doi: 10.3389/ebm.2025.10600. eCollection 2025. Exp Biol Med (Maywood). 2025. PMID: 40510242 Free PMC article.
References
-
- Shen L., Wu L.C., Sanlioglu S., Chen R., Mendoza A.R., Dangel A.W., Carroll M.C., Zipf W.B., Yu C.-Y.. Structure and genetics of the partially duplicated gene RP located immediately upstream of the complement C4A and the C4B genes in the HLA class III region. Molecular cloning, exon-intron structure, composite retroposon, and breakpoint of gene duplication. J. Biol. Chem. 1994; 269:8466–8476. - PubMed
-
- Wang H., Xing J., Grover D., Hedges D.J., Han K., Walker J.A., Batzer M.A.. SVA elements: a hominid-specific retroposon family. J. Mol. Biol. 2005; 354:994–1007. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
