

Comparative transcriptomics reveals human-specific cortical features
- PMID: 37824638
- PMCID: PMC10659116
- DOI: 10.1126/science.ade9516
Comparative transcriptomics reveals human-specific cortical features
Abstract
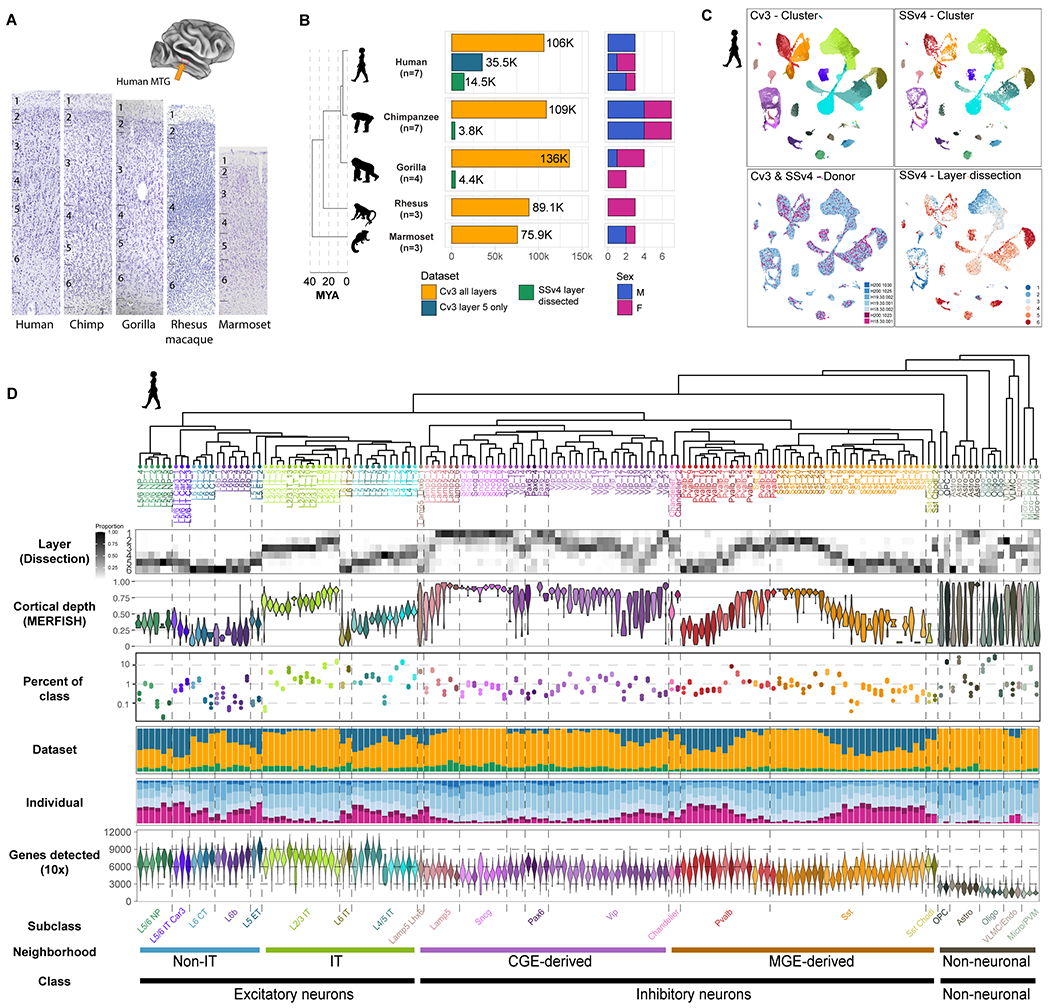
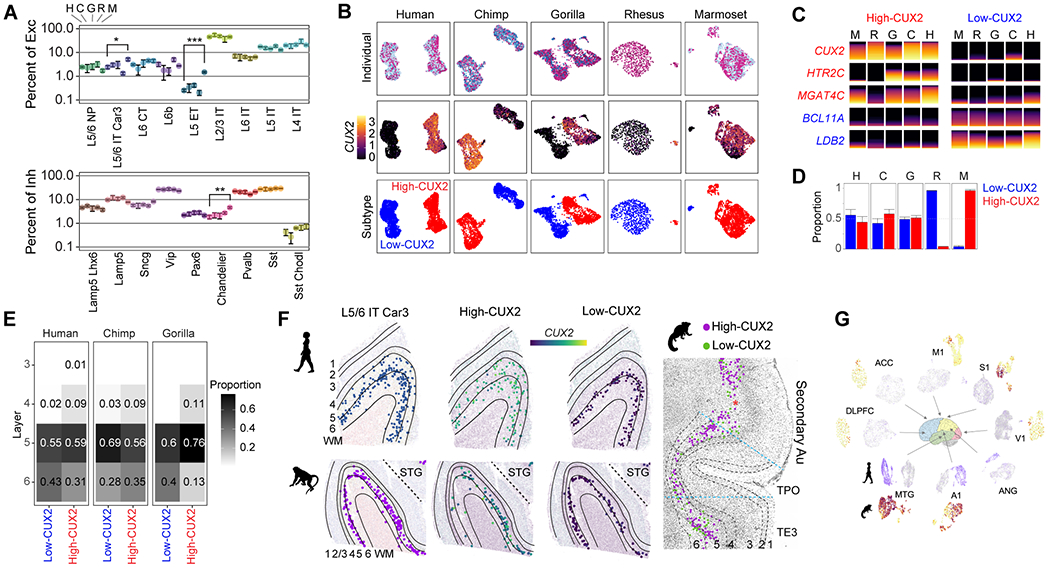
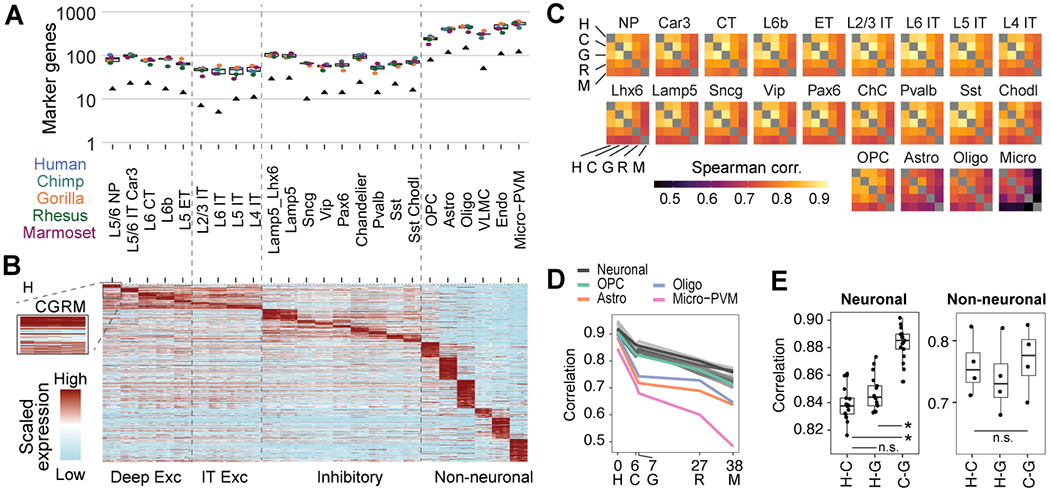
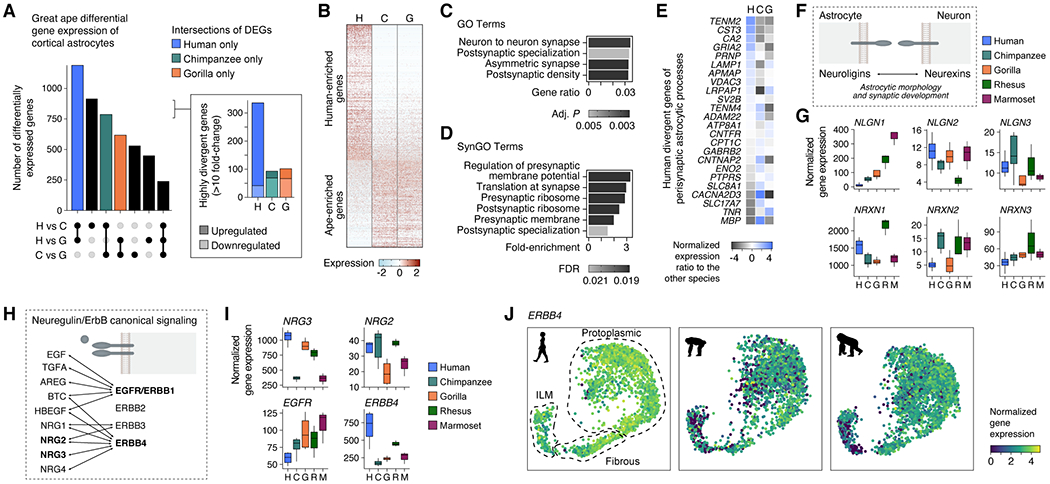
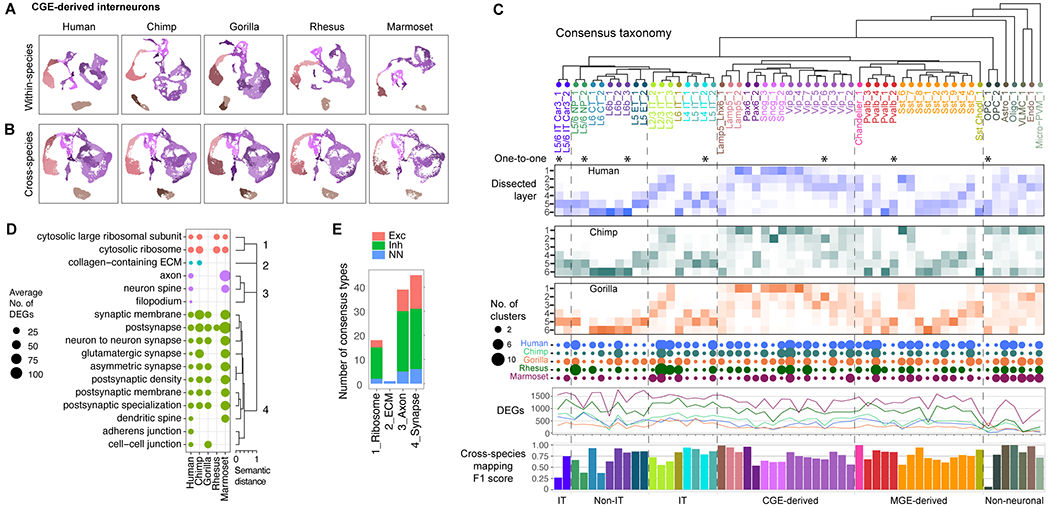
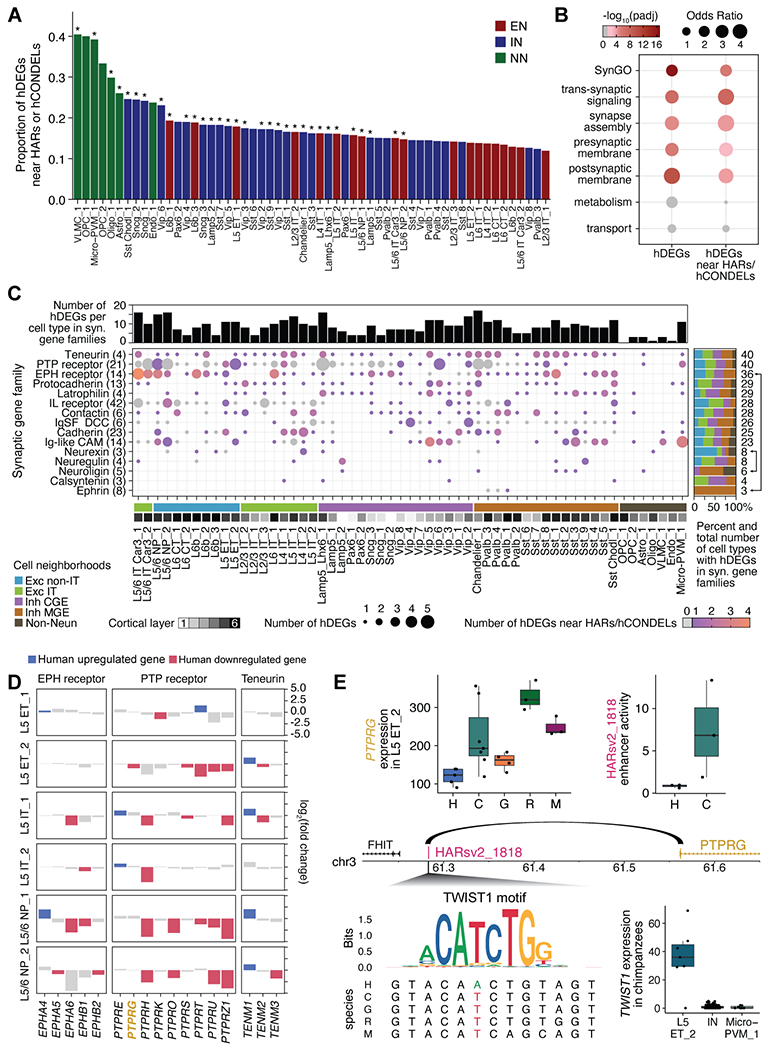
The cognitive abilities of humans are distinctive among primates, but their molecular and cellular substrates are poorly understood. We used comparative single-nucleus transcriptomics to analyze samples of the middle temporal gyrus (MTG) from adult humans, chimpanzees, gorillas, rhesus macaques, and common marmosets to understand human-specific features of the neocortex. Human, chimpanzee, and gorilla MTG showed highly similar cell-type composition and laminar organization as well as a large shift in proportions of deep-layer intratelencephalic-projecting neurons compared with macaque and marmoset MTG. Microglia, astrocytes, and oligodendrocytes had more-divergent expression across species compared with neurons or oligodendrocyte precursor cells, and neuronal expression diverged more rapidly on the human lineage. Only a few hundred genes showed human-specific patterning, suggesting that relatively few cellular and molecular changes distinctively define adult human cortical structure.
Conflict of interest statement
Figures
References
-
- Roumazeilles L, Eichert N, Bryant KL, Folloni D, Sallet J, Vijayakumar S, Foxley S, Tendler BC, Jbabdi S, Reveley C, Verhagen L, Dershowitz LB, Guthrie M, Flach E, Miller KL, Mars RB, Longitudinal connections and the organization of the temporal cortex in macaques, great apes, and humans. PLoS Biol. 18, e3000810 (2020). - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
