

Adenosine A2A receptors control generalization of contextual fear in rats
- PMID: 37828000
- PMCID: PMC10570294
- DOI: 10.1038/s41398-023-02613-0
Adenosine A2A receptors control generalization of contextual fear in rats
Abstract
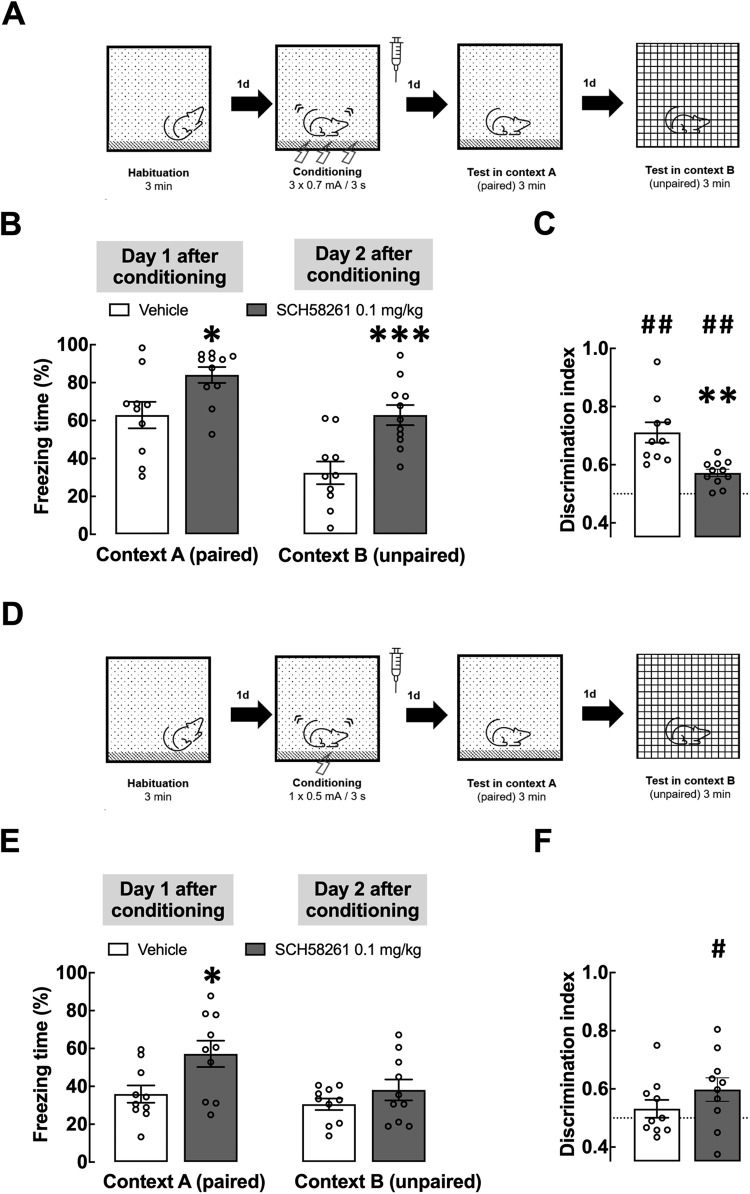
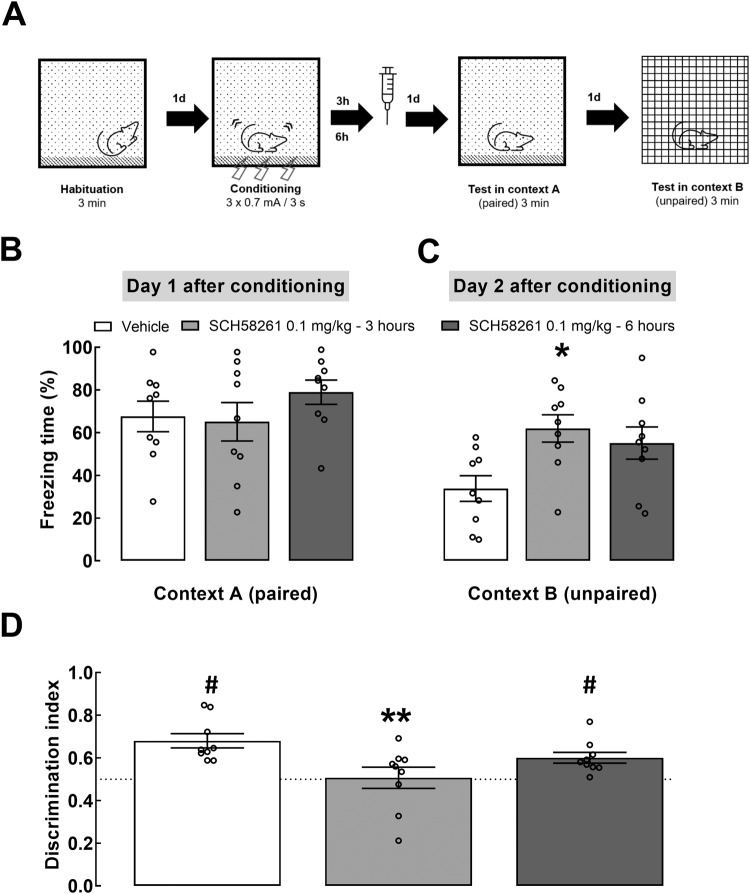
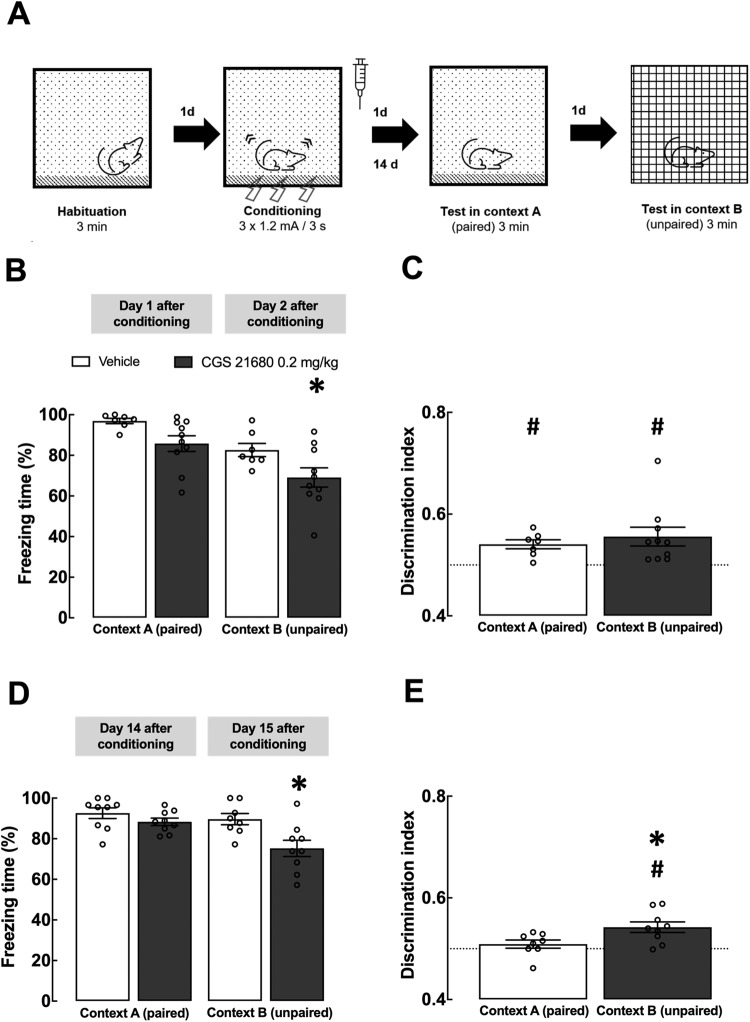
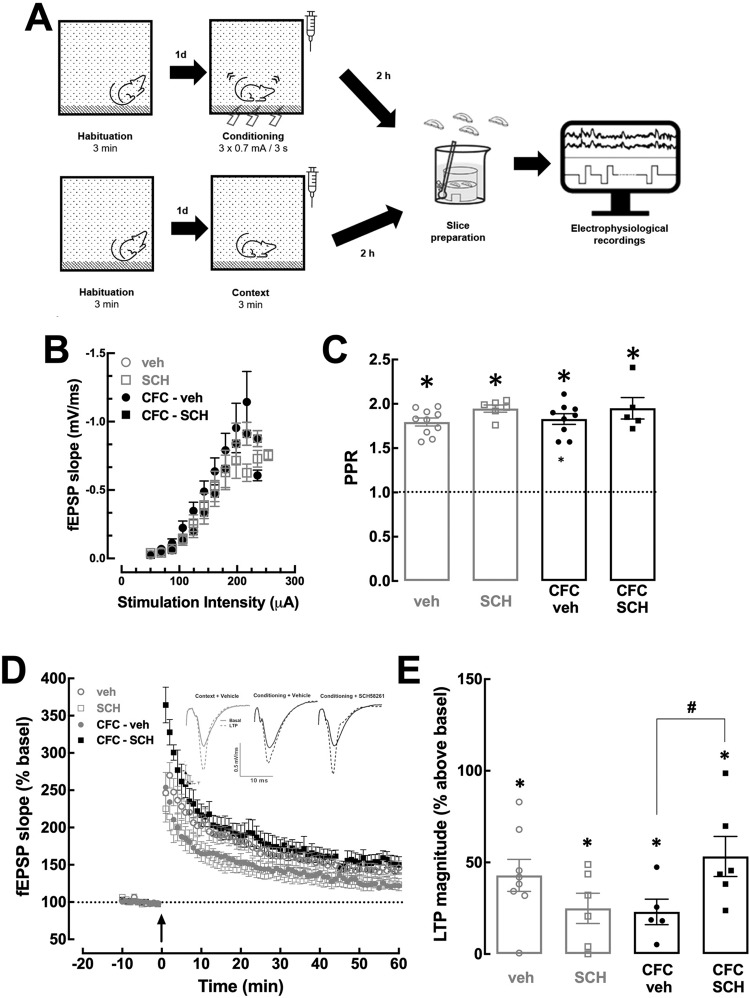
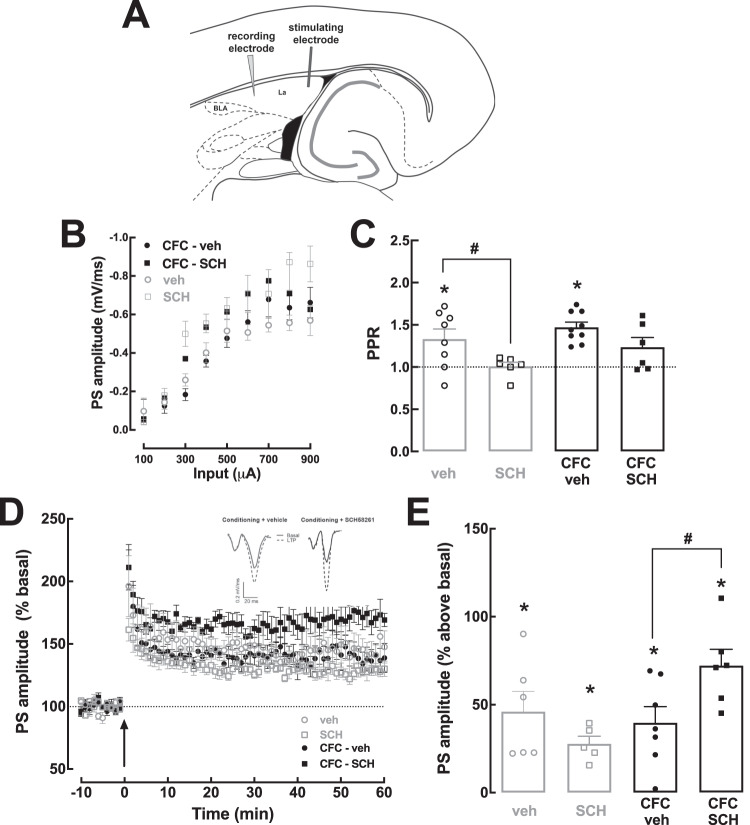
Fear learning is essential to survival, but traumatic events may lead to abnormal fear consolidation and overgeneralization, triggering fear responses in safe environments, as occurs in post-traumatic stress disorder (PTSD). Adenosine A2A receptors (A2AR) control emotional memory and fear conditioning, but it is not known if they affect the consolidation and generalization of fear, which was now investigated. We now report that A2AR blockade through systemic administration of the A2AR antagonist SCH58261 immediately after contextual fear conditioning (within the consolidation window), accelerated fear generalization. Conversely, A2AR activation with CGS21680 decreased fear generalization. Ex vivo electrophysiological recordings of field excitatory post-synaptic potentials (fEPSPs) in CA3-CA1 synapses and of population spikes in the lateral amygdala (LA), showed that the effect of SCH58261 is associated with a reversion of fear conditioning-induced decrease of long-term potentiation (LTP) in the dorsal hippocampus (DH) and with increased amplitude of LA LTP in conditioned animals. These data suggest that A2AR are engaged during contextual fear consolidation, controlling long-term potentiation mechanisms in both DH and LA during fear consolidation, impacting on fear generalization; this supports targeting A2AR during fear consolidation to control aberrant fear processing in PTSD and other fear-related disorders.
© 2023. Springer Nature Limited.
Conflict of interest statement
RAC is a scientific consultant for the Institute for Scientific Information on Coffee (ISIC). All other authors declare no competing financial interests.
Figures

References
-
- Dunsmoor JE, Paz R. Fear generalization and anxiety: behavioral and neural mechanisms. Biol Psychiatry. 2015;78:336–43. - PubMed
-
- Izquierdo I, Furini CR, Myskiw JC. Fear memory. Physiol Rev. 2016;96:695–750. - PubMed
-
- Holland PC, Bouton ME. Hippocampus and context in classical conditioning. Curr Opin Neurobiol. 1999;9:195–202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- LCF/PR/HP17/52190001/"la Caixa" Foundation (Caixa Foundation)
- POCI-01-0145-FEDER-031274/Ministry of Education and Science | Fundação para a Ciência e a Tecnologia (Portuguese Science and Technology Foundation)
- UIDB/04539/2020/Ministry of Education and Science | Fundação para a Ciência e a Tecnologia (Portuguese Science and Technology Foundation)
LinkOut - more resources
Full Text Sources
Miscellaneous
