

Axon-Autonomous Effects of the Amyloid Precursor Protein Intracellular Domain (AICD) on Kinase Signaling and Fast Axonal Transport
- PMID: 37830617
- PMCID: PMC10572015
- DOI: 10.3390/cells12192403
Axon-Autonomous Effects of the Amyloid Precursor Protein Intracellular Domain (AICD) on Kinase Signaling and Fast Axonal Transport
Abstract
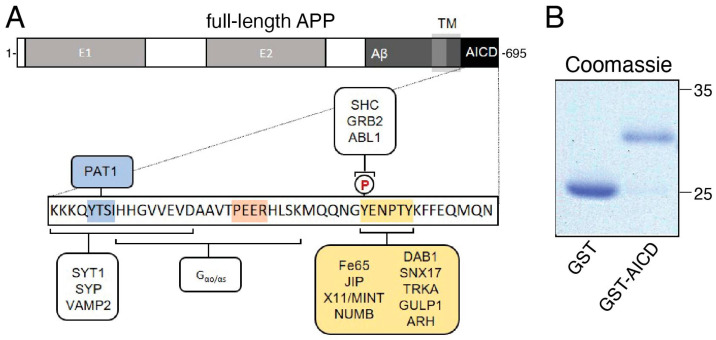
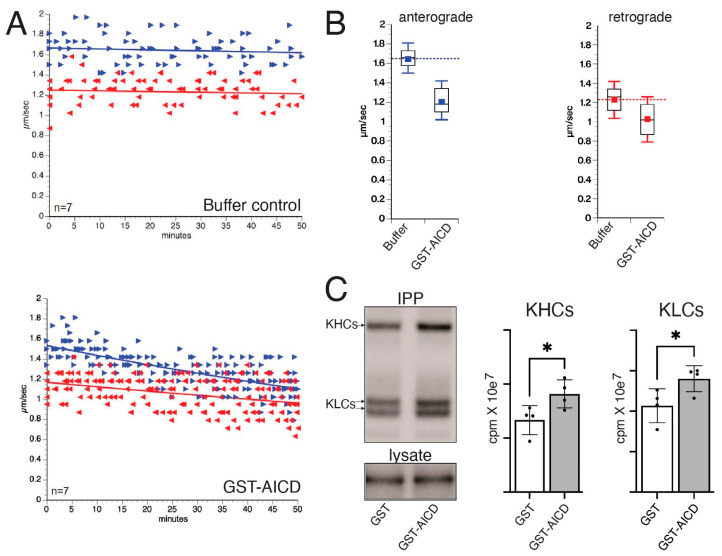
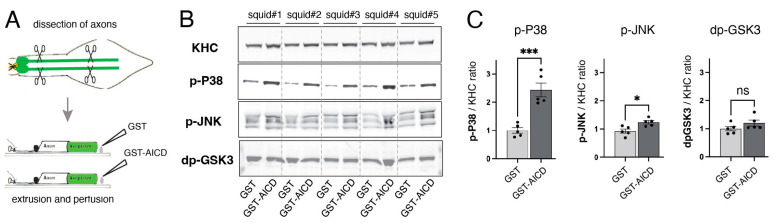
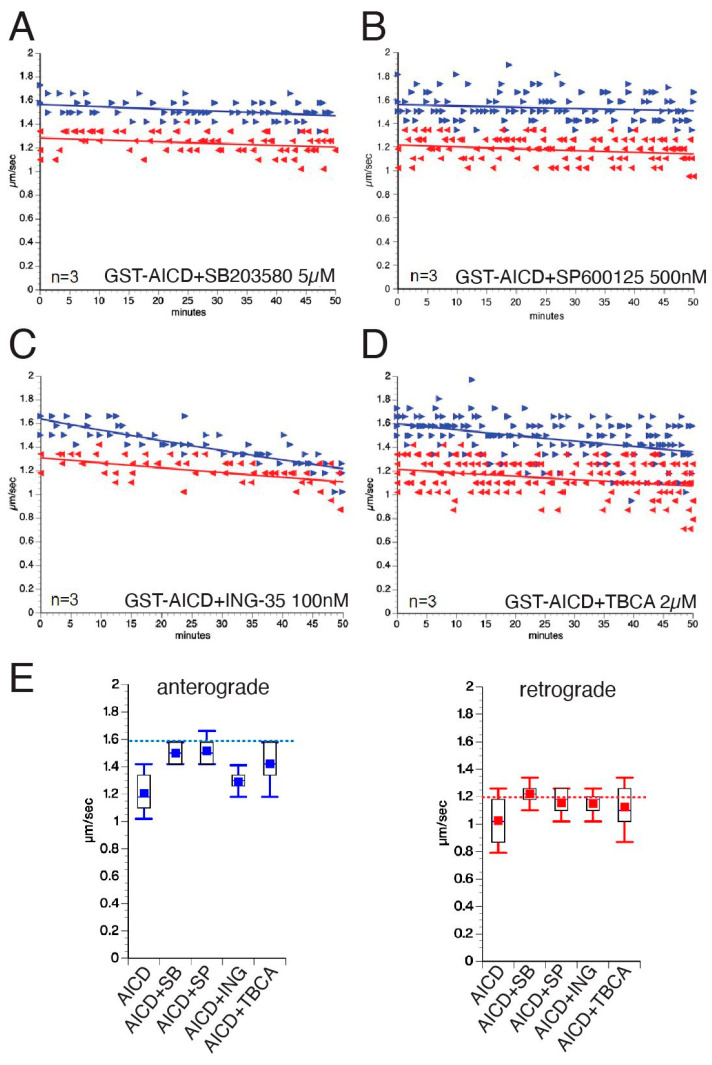
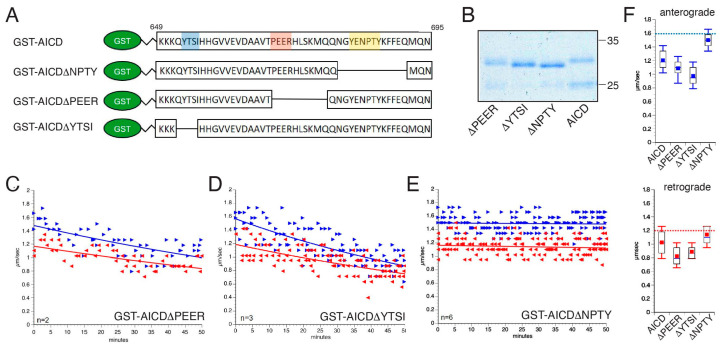
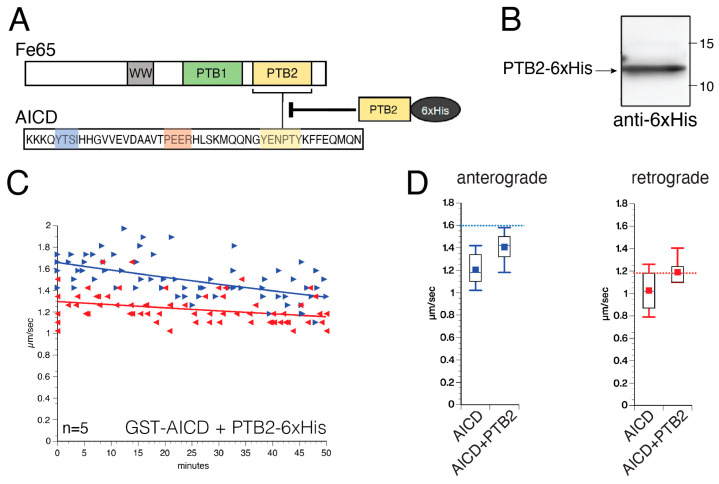
The amyloid precursor protein (APP) is a key molecular component of Alzheimer's disease (AD) pathogenesis. Proteolytic APP processing generates various cleavage products, including extracellular amyloid beta (Aβ) and the cytoplasmic APP intracellular domain (AICD). Although the role of AICD in the activation of kinase signaling pathways is well established in the context of full-length APP, little is known about intracellular effects of the AICD fragment, particularly within discrete neuronal compartments. Deficits in fast axonal transport (FAT) and axonopathy documented in AD-affected neurons prompted us to evaluate potential axon-autonomous effects of the AICD fragment for the first time. Vesicle motility assays using the isolated squid axoplasm preparation revealed inhibition of FAT by AICD. Biochemical experiments linked this effect to aberrant activation of selected axonal kinases and heightened phosphorylation of the anterograde motor protein conventional kinesin, consistent with precedents showing phosphorylation-dependent regulation of motors proteins powering FAT. Pharmacological inhibitors of these kinases alleviated the AICD inhibitory effect on FAT. Deletion experiments indicated this effect requires a sequence encompassing the NPTY motif in AICD and interacting axonal proteins containing a phosphotyrosine-binding domain. Collectively, these results provide a proof of principle for axon-specific effects of AICD, further suggesting a potential mechanistic framework linking alterations in APP processing, FAT deficits, and axonal pathology in AD.
Keywords: AICD; APP; Alzheimer’s disease; NPTY; kinases.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Morfini G.A., You Y.-M., Pollema S.L., Kaminska A., Liu K., Yoshioka K., Björkblom B., Coffey E.T., Bagnato C., Han D., et al. Pathogenic huntingtin inhibits fast axonal transport by activating JNK3 and phosphorylating kinesin. Nat. Neurosci. 2009;12:864–871. doi: 10.1038/nn.2346. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
