

IL-31-dependent neurogenic inflammation restrains cutaneous type 2 immune response in allergic dermatitis
- PMID: 37831760
- PMCID: PMC10890830
- DOI: 10.1126/sciimmunol.abi6887
IL-31-dependent neurogenic inflammation restrains cutaneous type 2 immune response in allergic dermatitis
Abstract
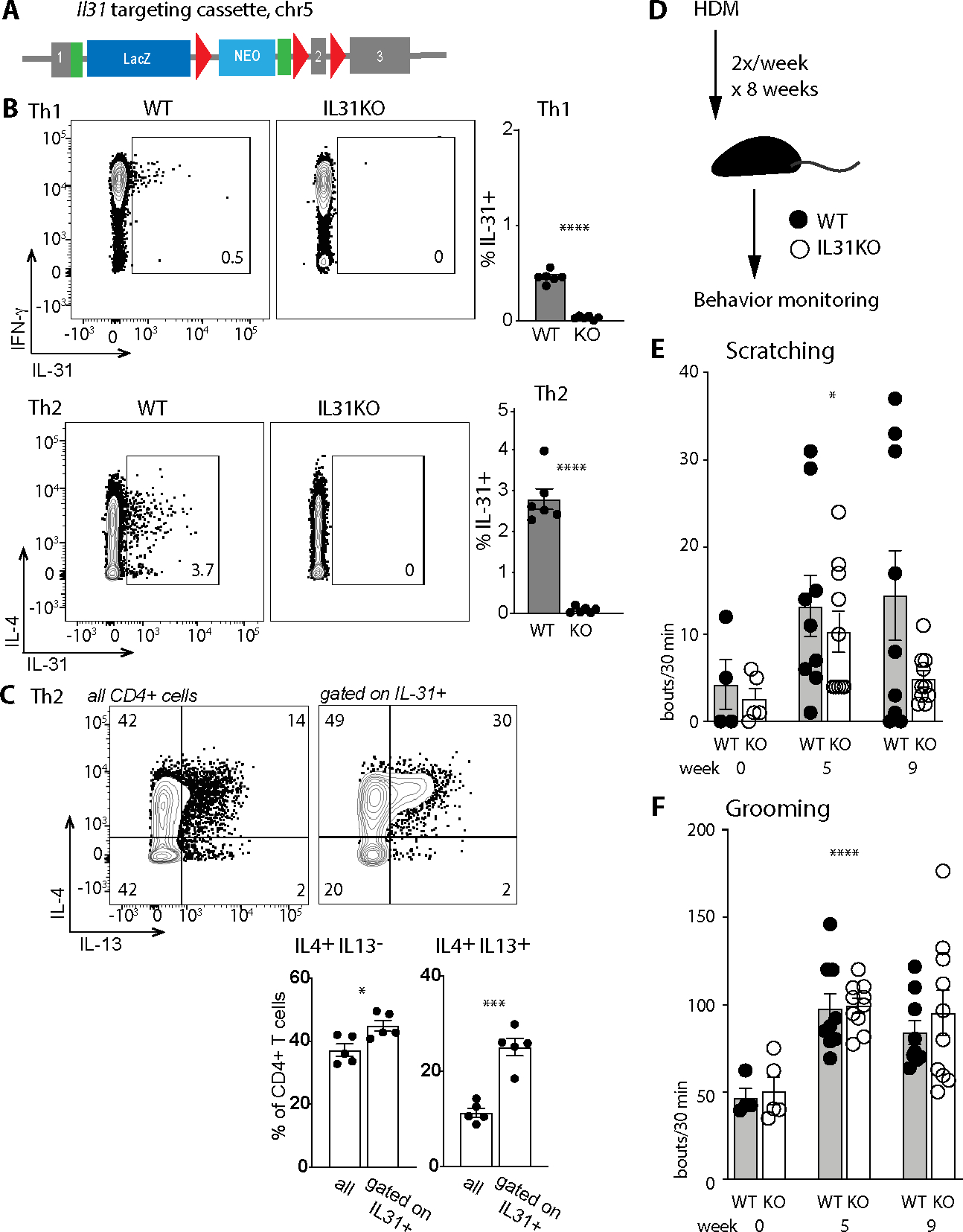
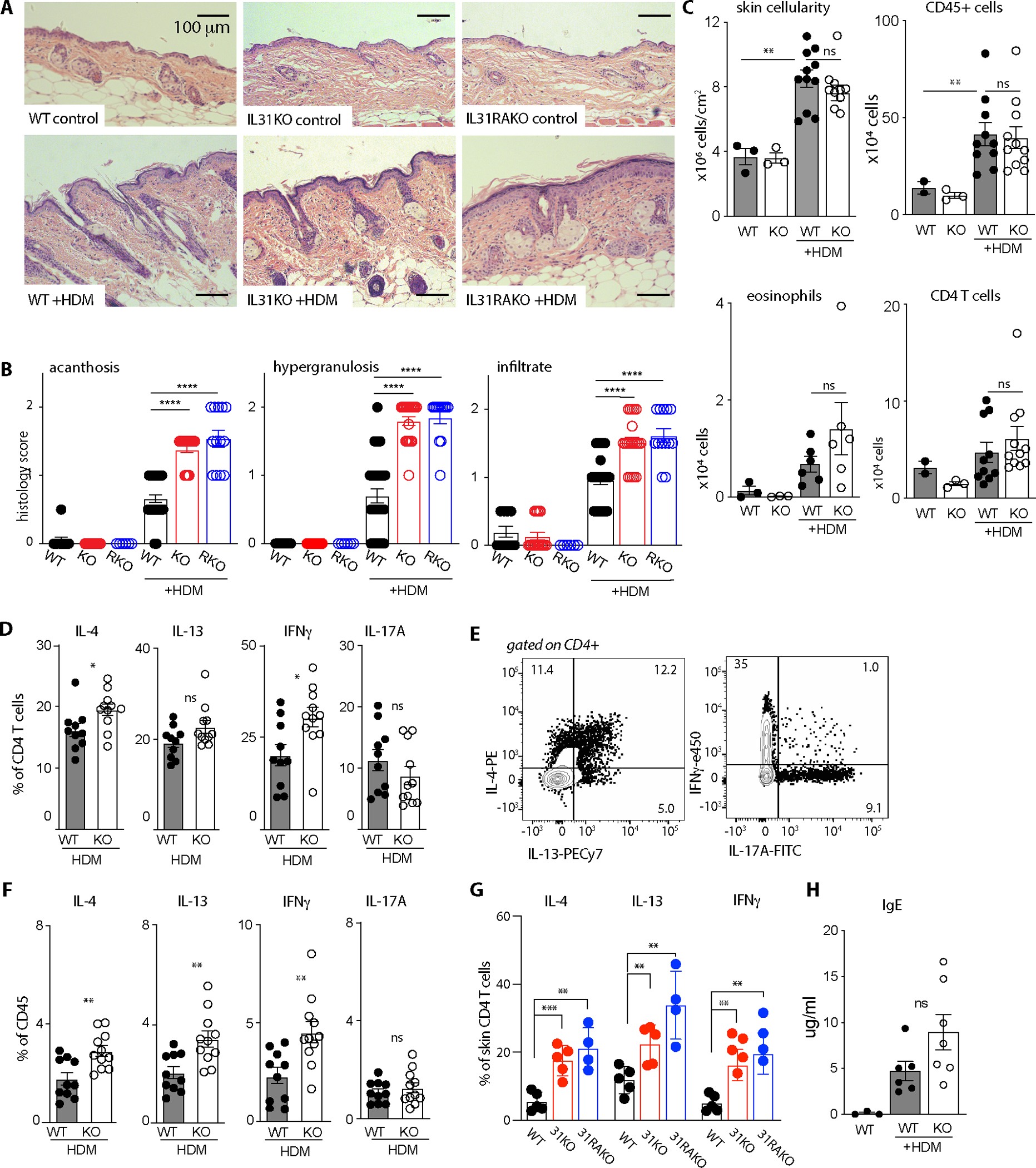
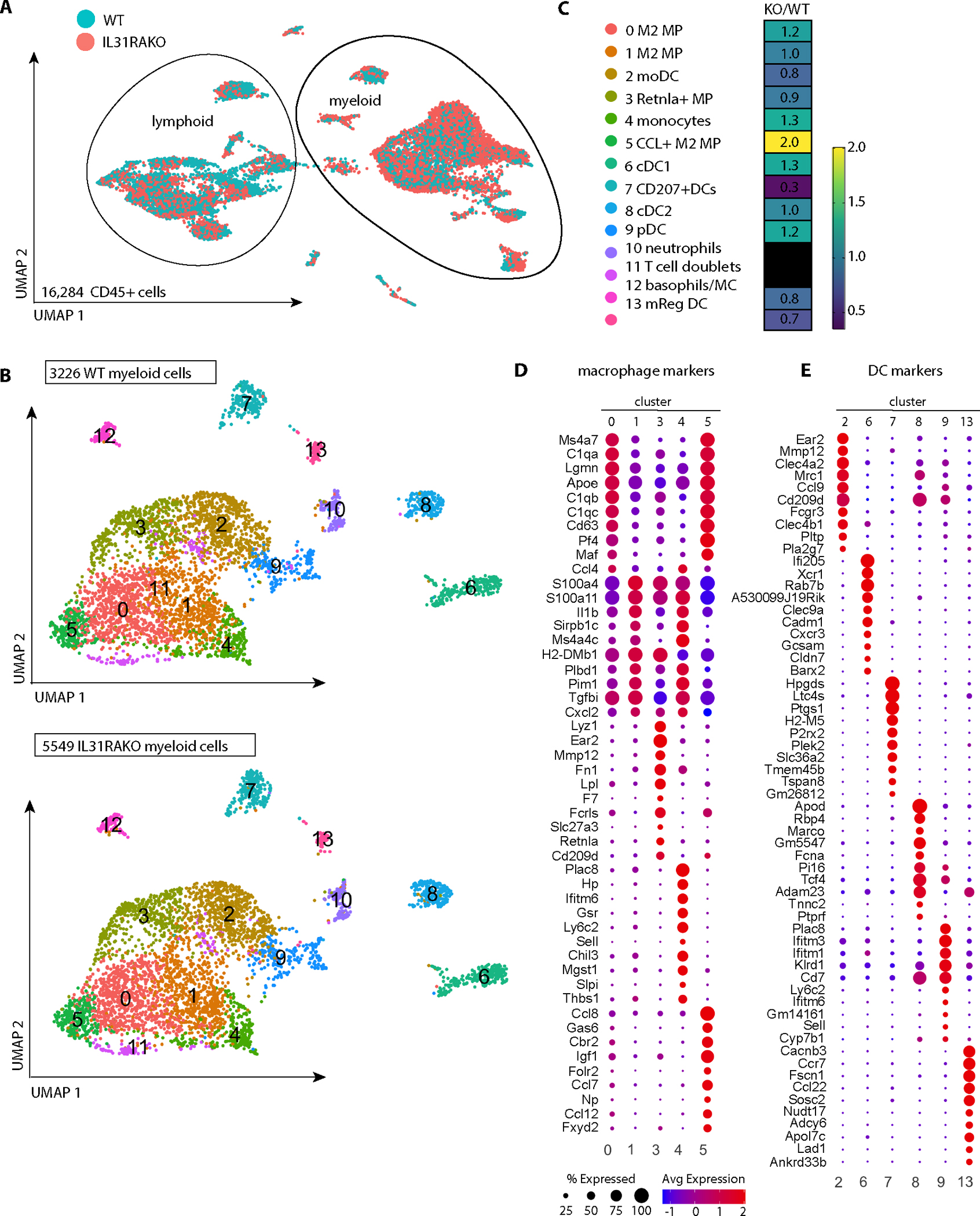
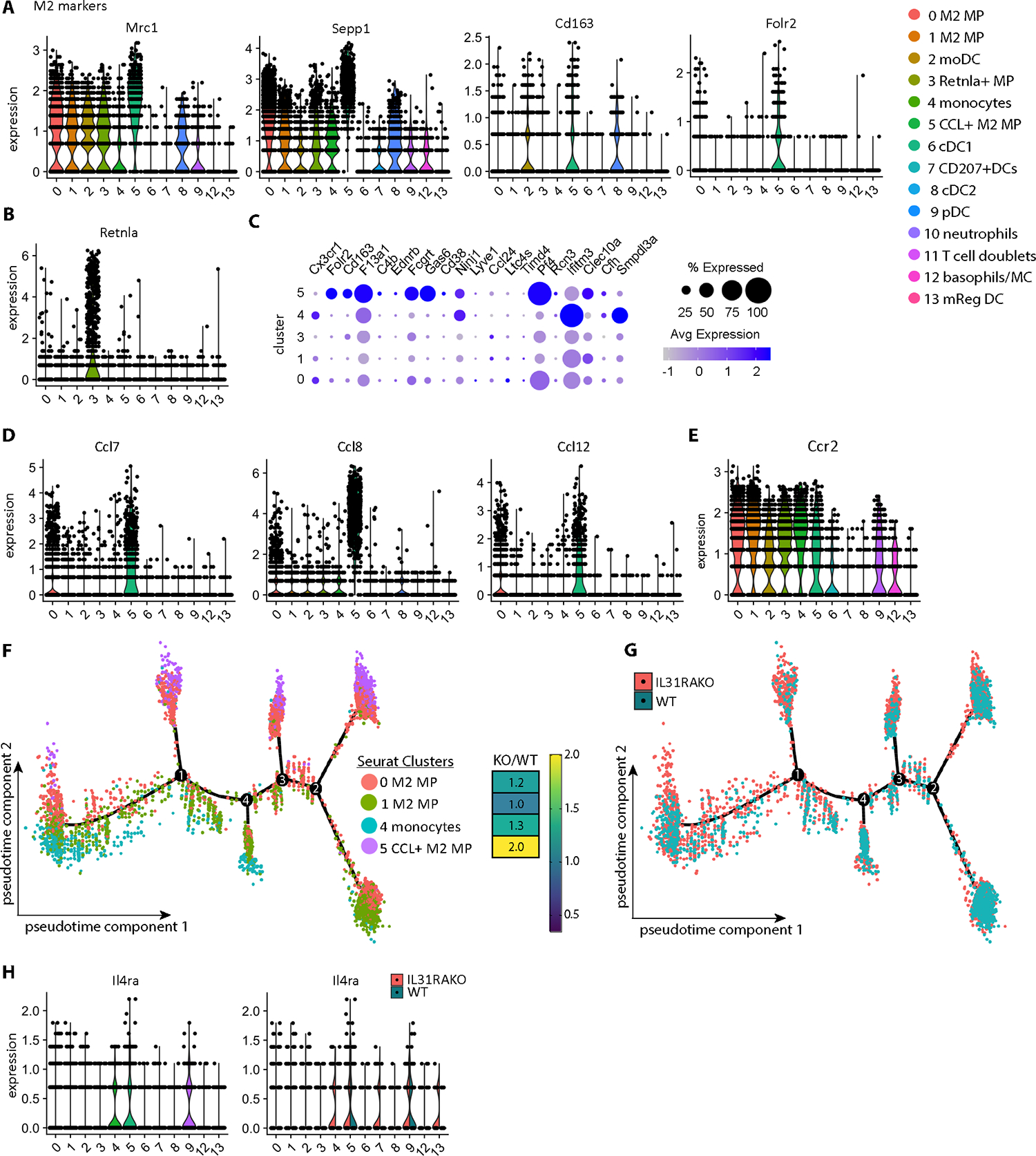
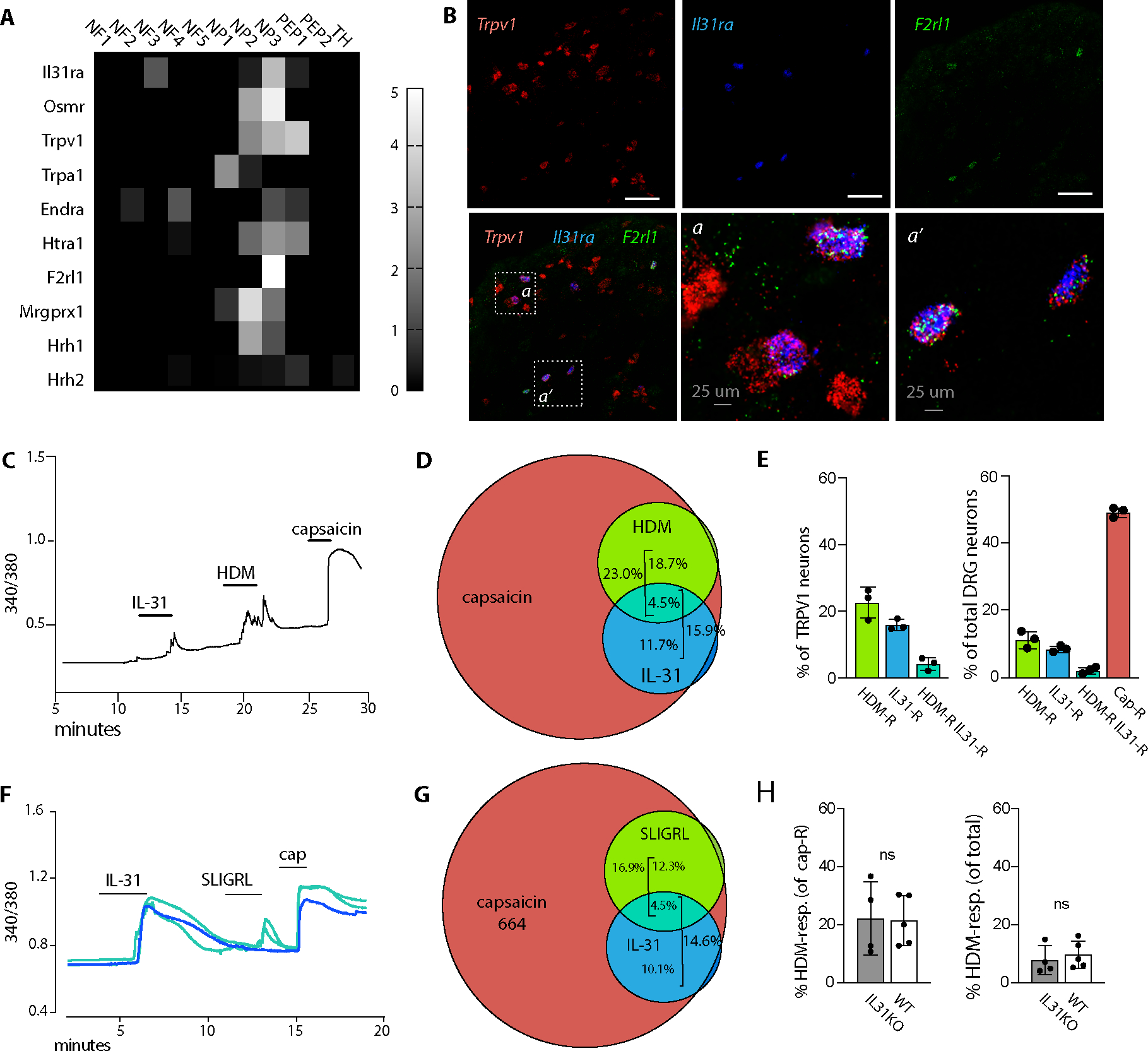
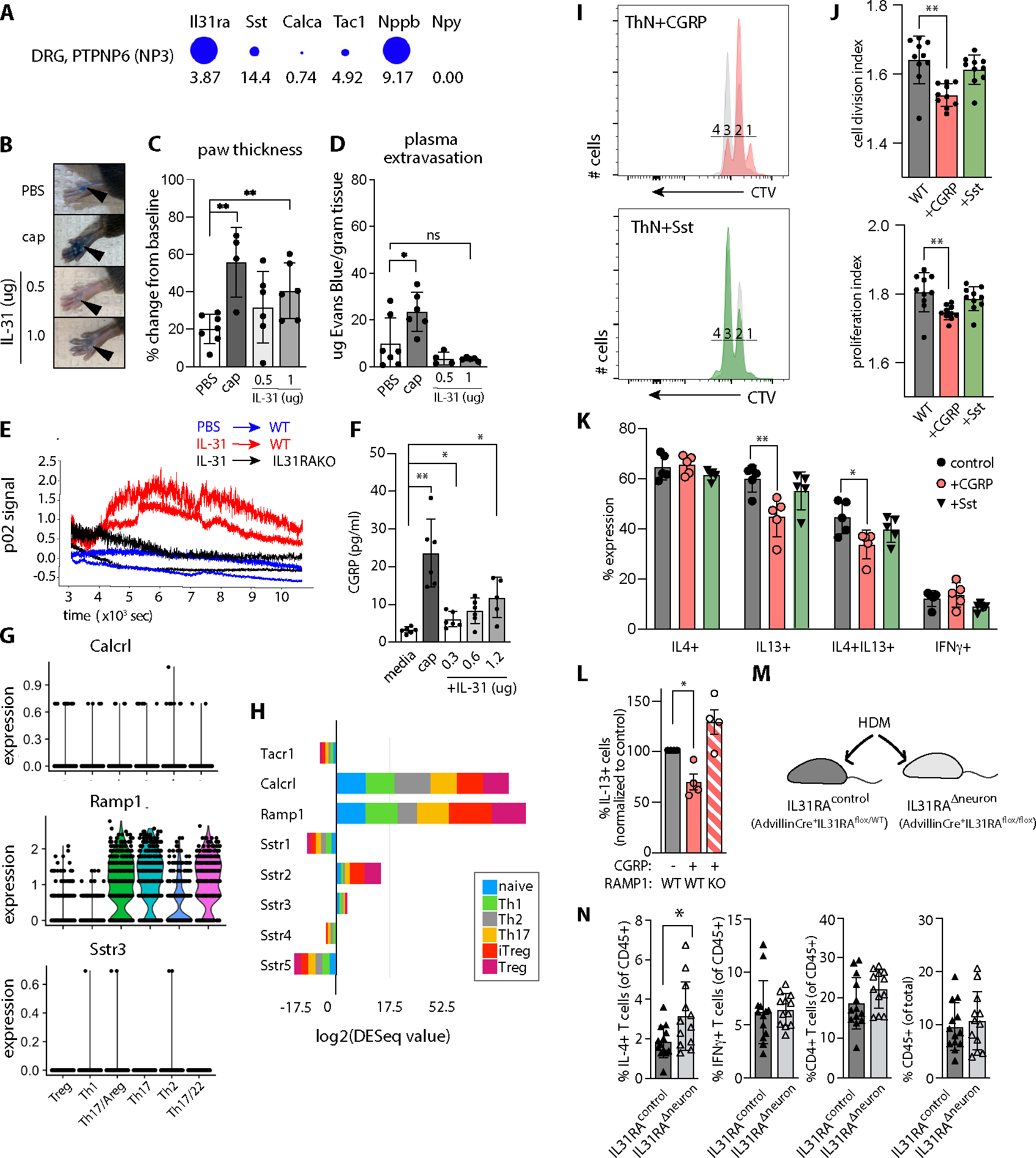
Despite robust literature associating IL-31 with pruritic inflammatory skin diseases, its influence on cutaneous inflammation and the interplay between inflammatory and neurosensory pathways remain unmapped. Here, we examined the consequences of disrupting Il31 and its receptor Il31ra in a mouse model of house dust mite (HDM)-induced allergic dermatitis. Il31-deficient mice displayed a deficit in HDM dermatitis-associated scratching, consistent with its well-established role as a pruritogen. In contrast, Il31 deficiency increased the number and proportion of cutaneous type 2 cytokine-producing CD4+ T cells and serum IgE in response to HDM. Furthermore, Il4ra+ monocytes and macrophages capable of fueling a feedforward type 2 inflammatory loop were selectively enriched in Il31ra-deficient HDM dermatitis skin. Thus, IL-31 is not strictly a proinflammatory cytokine but rather an immunoregulatory factor that limits the magnitude of type 2 inflammatory responses in skin. Our data support a model wherein IL-31 activation of IL31RA+ pruritoceptors triggers release of calcitonin gene-related protein (CGRP), which can mediate neurogenic inflammation, inhibit CD4+ T cell proliferation, and reduce T cell production of the type 2 cytokine IL-13. Together, these results illustrate a previously unrecognized neuroimmune pathway that constrains type 2 tissue inflammation in the setting of chronic cutaneous allergen exposure and may explain paradoxical dermatitis flares in atopic patients treated with anti-IL31RA therapy.
Conflict of interest statement
Figures
References
-
- S. R. Dillon et al. , Interleukin 31, a cytokine produced by activated T cells, induces dermatitis in mice. Nat. Immunol. 5, 752–760 (2004). - PubMed
-
- Kabashima K, Matsumura T, Komazaki H, Kawashima M, Nemolizumab JPSG, Trial of Nemolizumab and Topical Agents for Atopic Dermatitis with Pruritus. N. Engl. J. Med. 383, 141–150 (2020). - PubMed
-
- Stander S et al. , Trial of Nemolizumab in Moderate-to-Severe Prurigo Nodularis. N. Engl. J. Med. 382, 706–716 (2020). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
