

Glial-derived mitochondrial signals affect neuronal proteostasis and aging
- PMID: 37831769
- PMCID: PMC10575585
- DOI: 10.1126/sciadv.adi1411
Glial-derived mitochondrial signals affect neuronal proteostasis and aging
Abstract
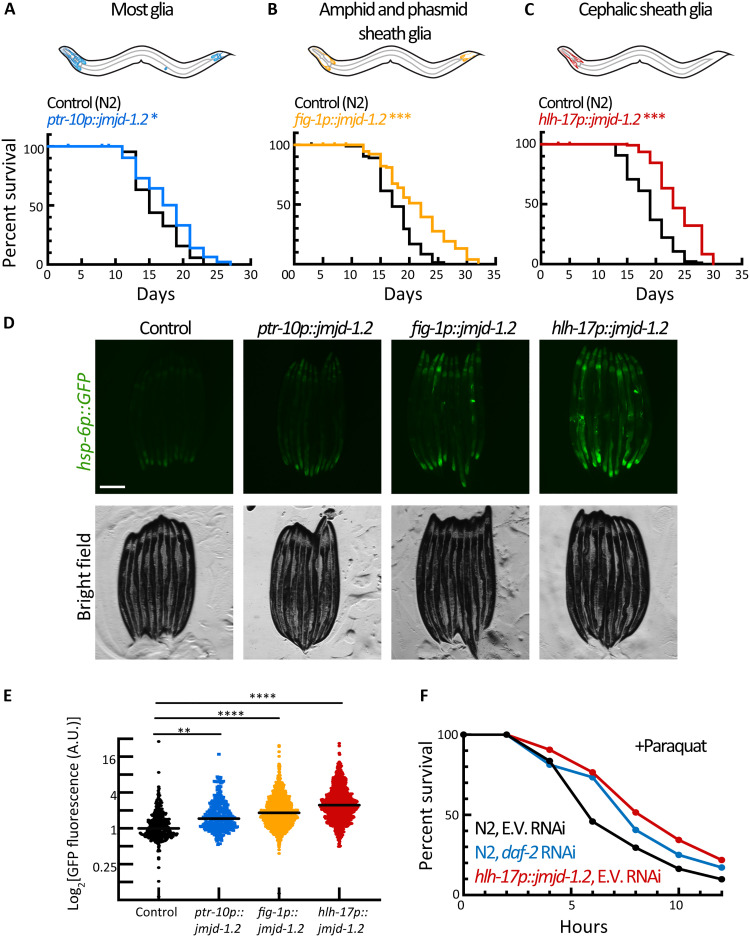
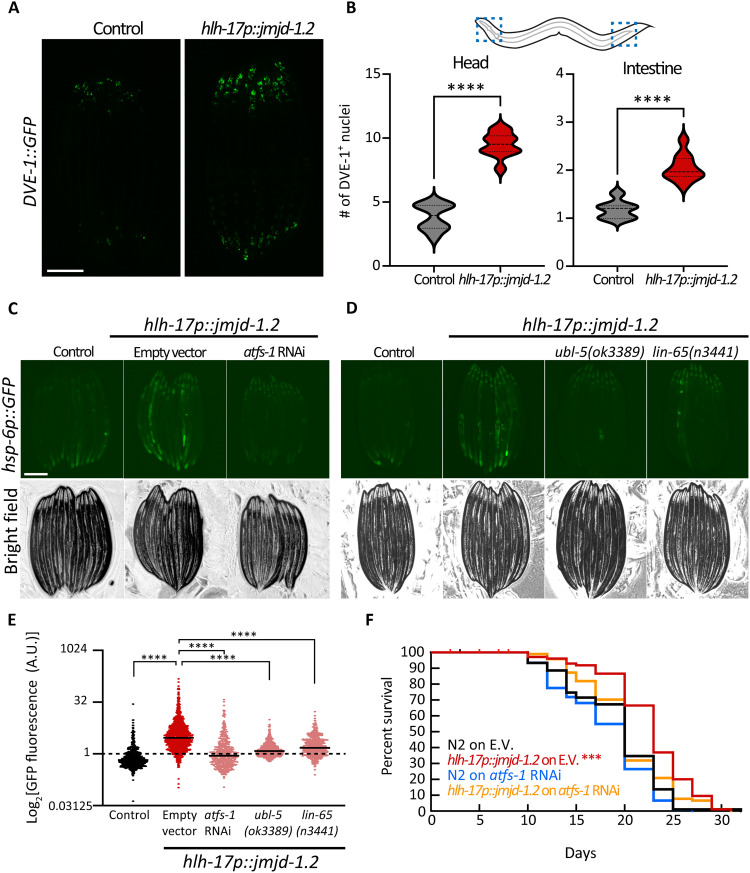
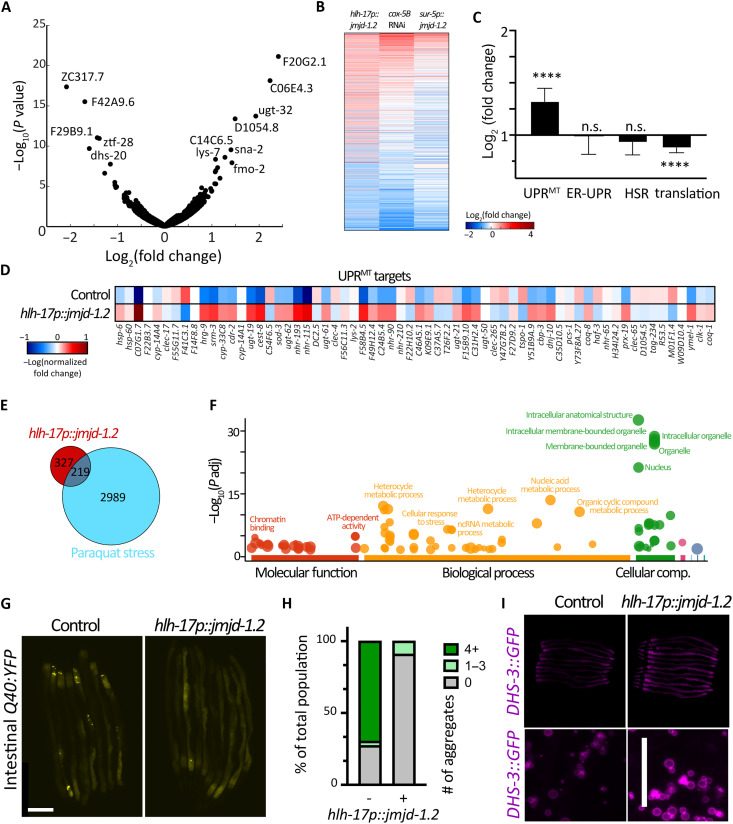
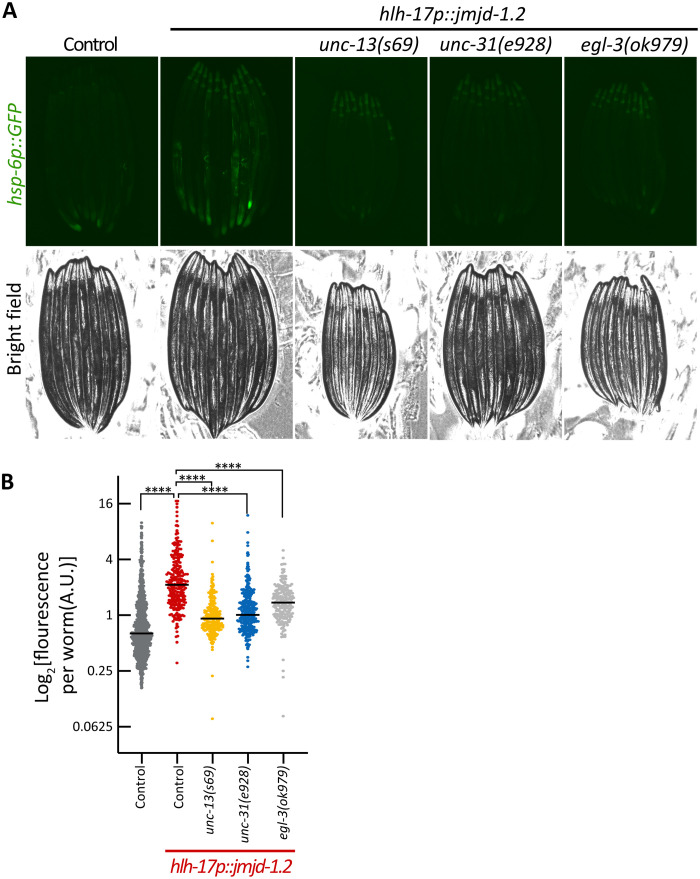
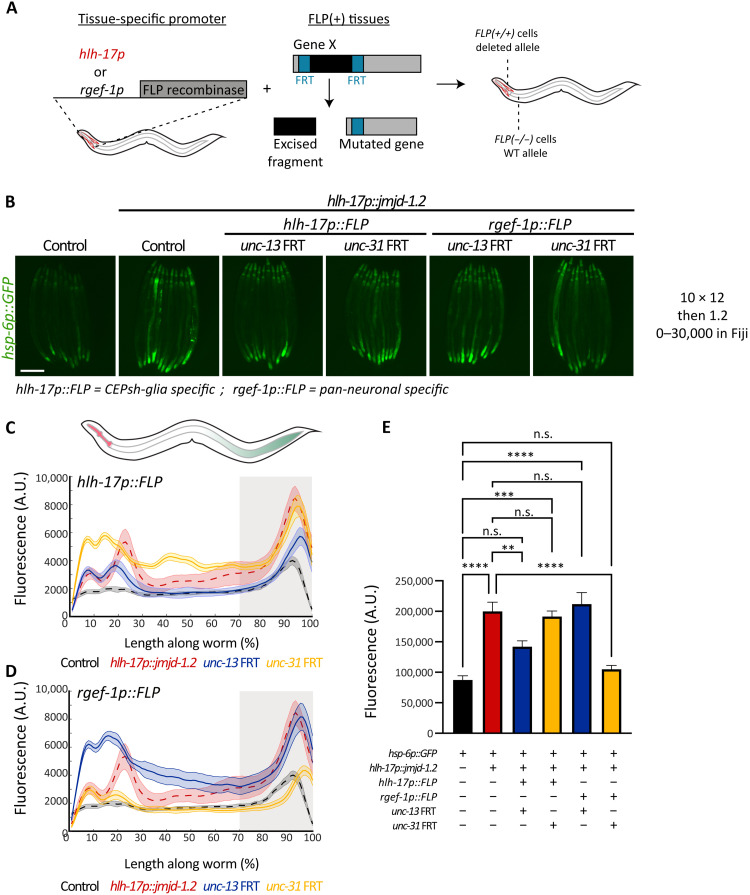
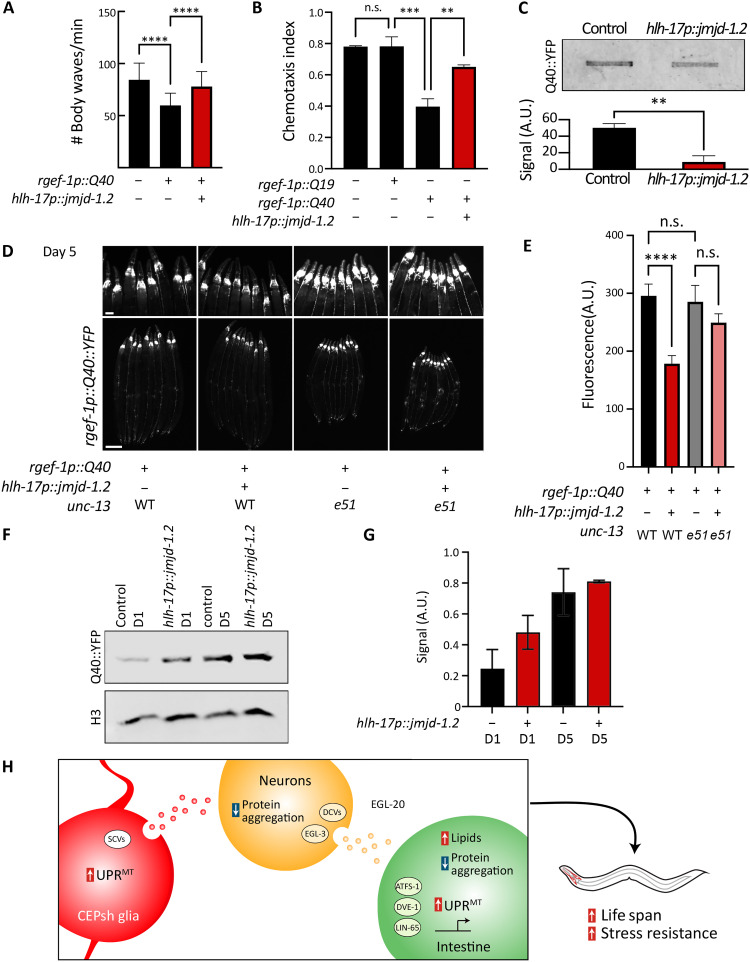
The nervous system plays a critical role in maintaining whole-organism homeostasis; neurons experiencing mitochondrial stress can coordinate the induction of protective cellular pathways, such as the mitochondrial unfolded protein response (UPRMT), between tissues. However, these studies largely ignored nonneuronal cells of the nervous system. Here, we found that UPRMT activation in four astrocyte-like glial cells in the nematode, Caenorhabditis elegans, can promote protein homeostasis by alleviating protein aggregation in neurons. Unexpectedly, we find that glial cells use small clear vesicles (SCVs) to signal to neurons, which then relay the signal to the periphery using dense-core vesicles (DCVs). This work underlines the importance of glia in establishing and regulating protein homeostasis within the nervous system, which can then affect neuron-mediated effects in organismal homeostasis and longevity.
Figures
Update of
-
Glial-derived mitochondrial signals impact neuronal proteostasis and aging.bioRxiv [Preprint]. 2023 Aug 8:2023.07.20.549924. doi: 10.1101/2023.07.20.549924. bioRxiv. 2023. Update in: Sci Adv. 2023 Oct 13;9(41):eadi1411. doi: 10.1126/sciadv.adi1411. PMID: 37609253 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
