

Sen1 architecture: RNA-DNA hybrid resolution, autoregulation, and insights into SETX inactivation in AOA2
- PMID: 37832548
- PMCID: PMC10629462
- DOI: 10.1016/j.molcel.2023.09.024
Sen1 architecture: RNA-DNA hybrid resolution, autoregulation, and insights into SETX inactivation in AOA2
Abstract
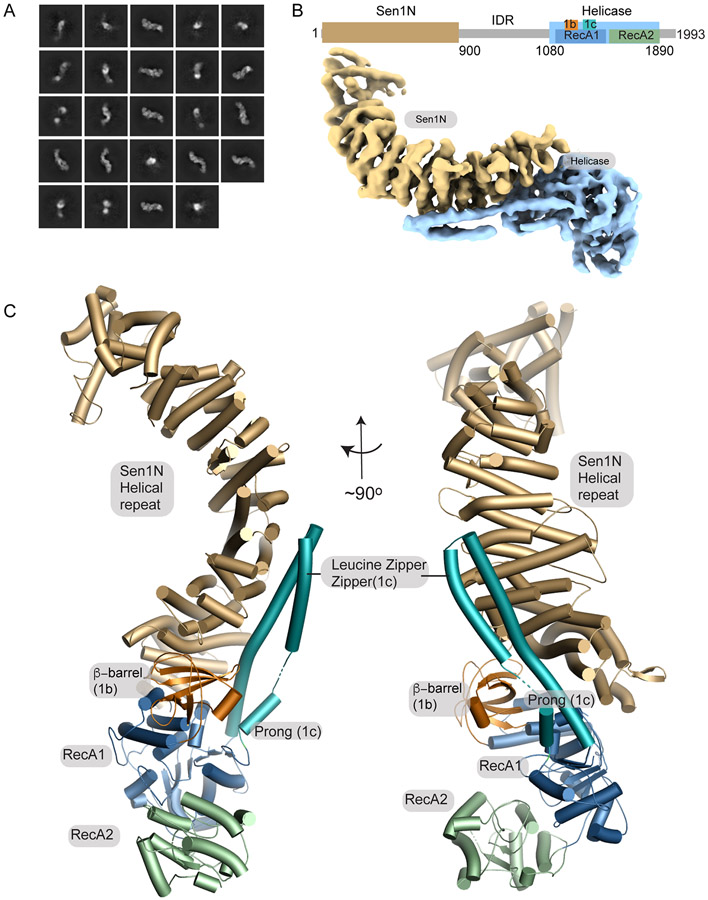
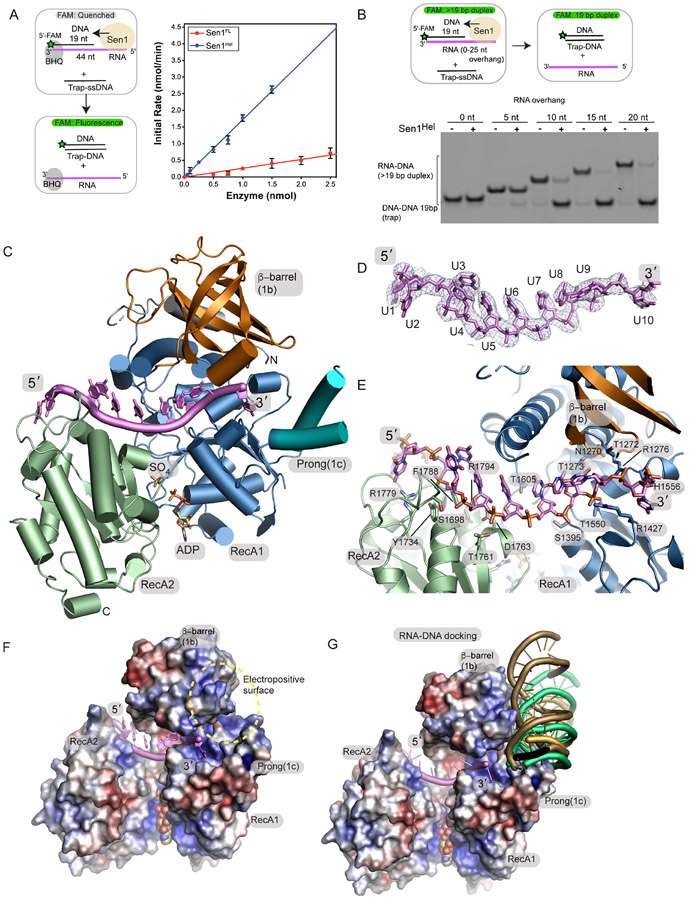
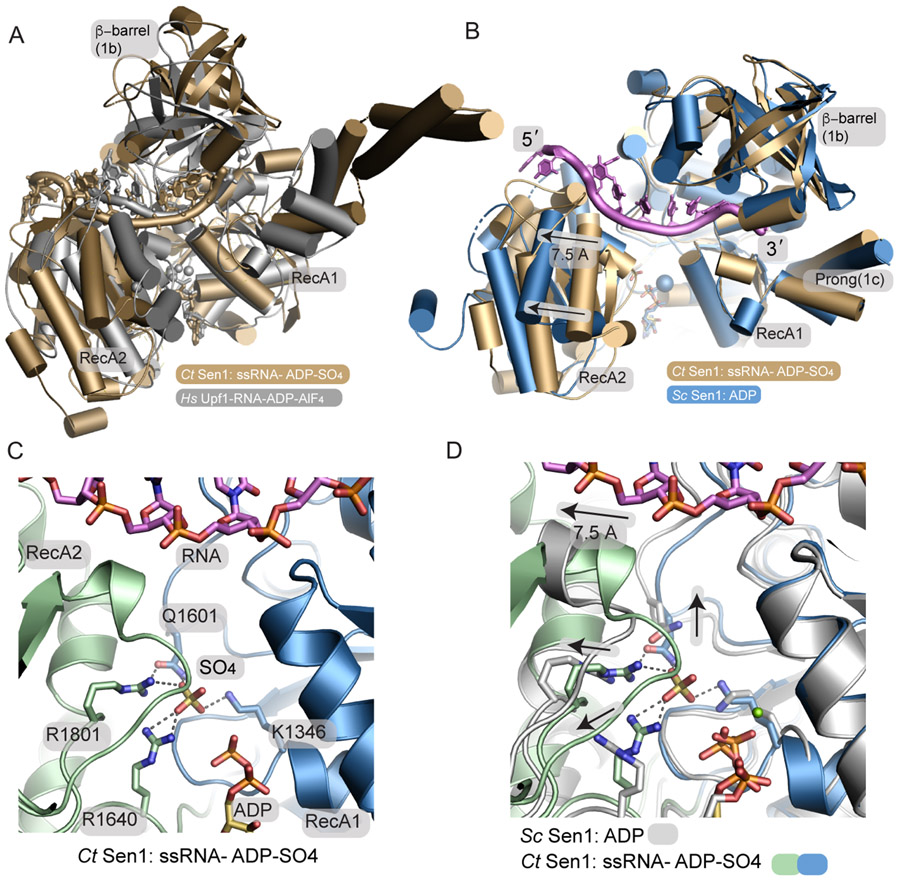
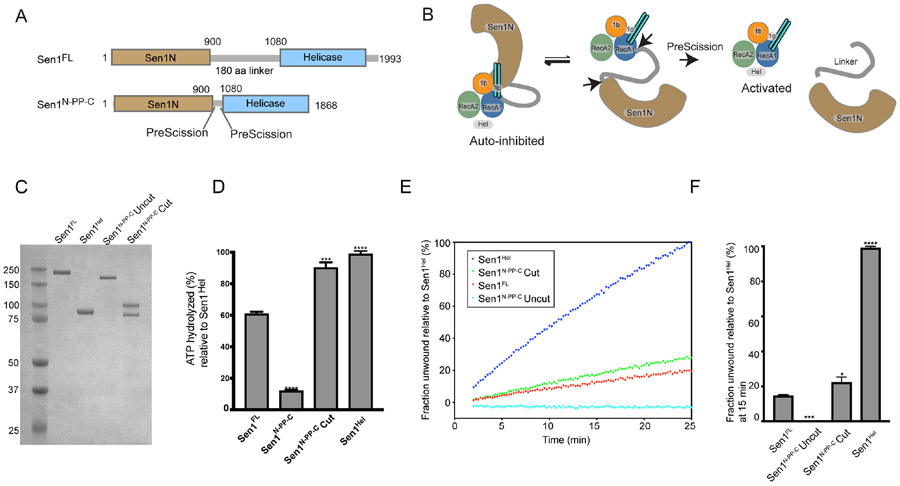
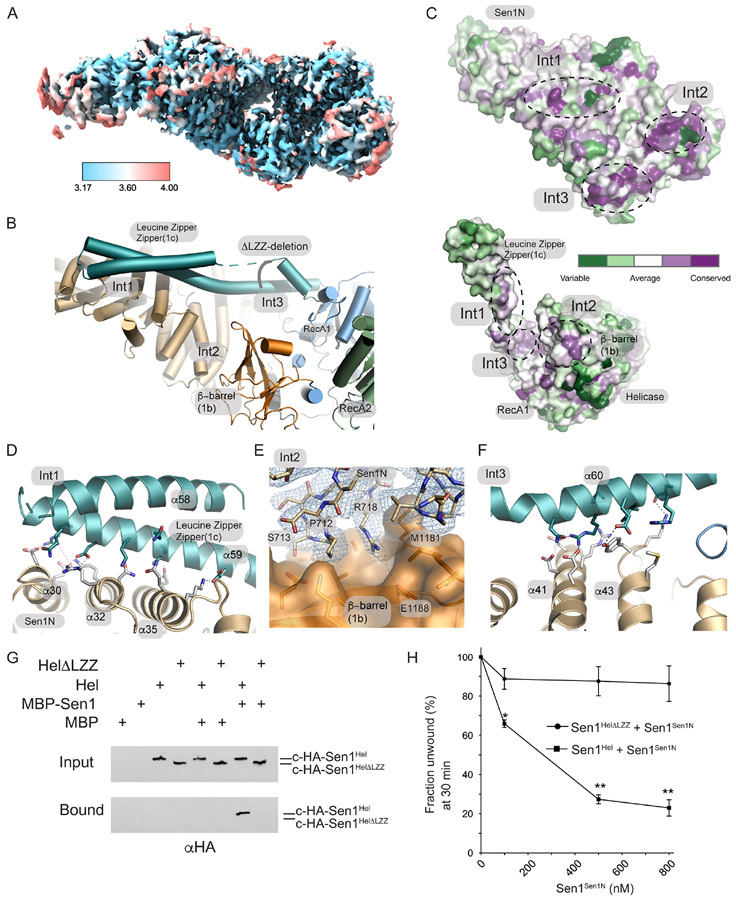
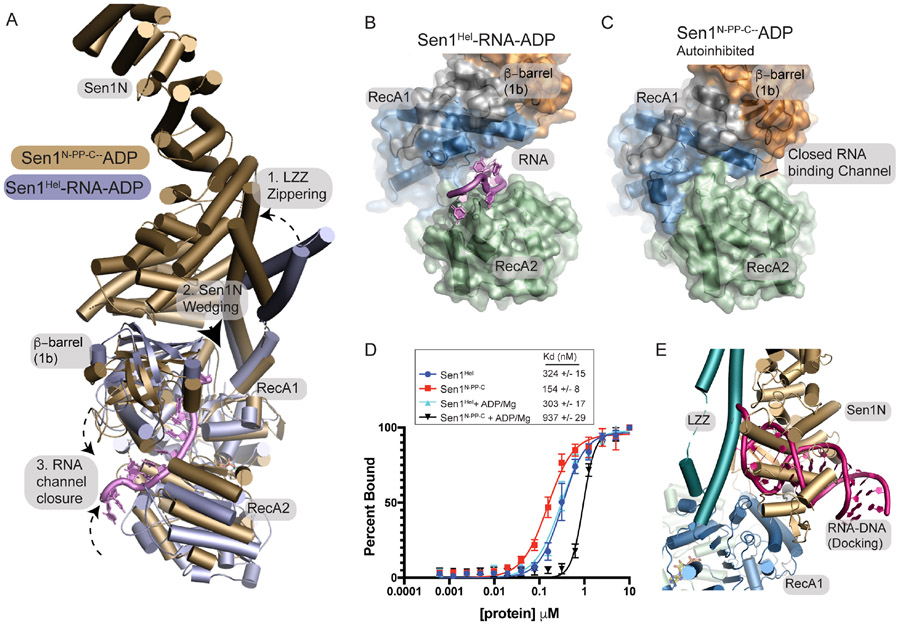
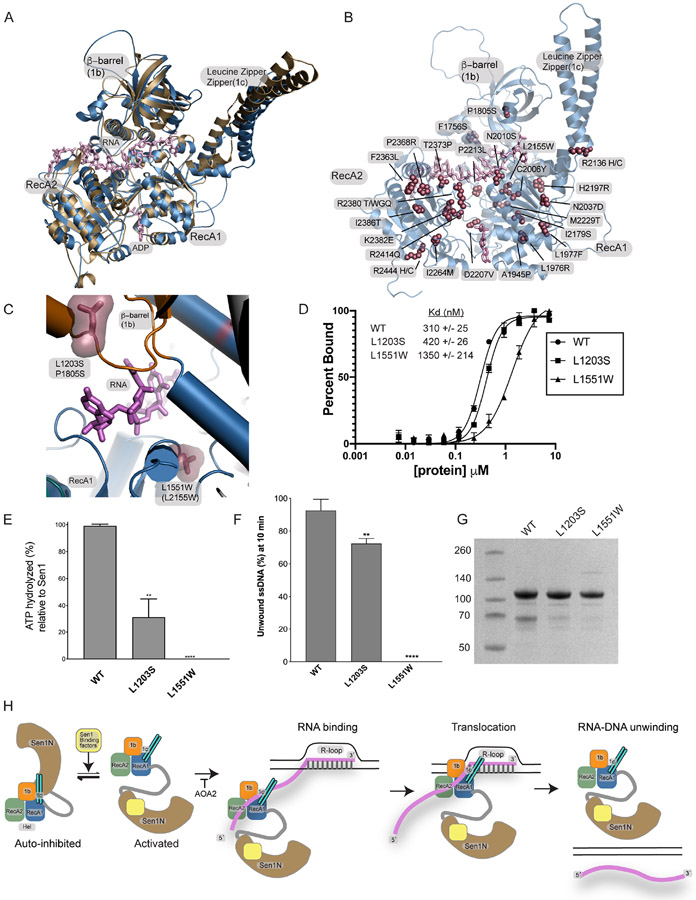
The senataxin (SETX, Sen1 in yeasts) RNA-DNA hybrid resolving helicase regulates multiple nuclear transactions, including DNA replication, transcription, and DNA repair, but the molecular basis for Sen1 activities is ill defined. Here, Sen1 cryoelectron microscopy (cryo-EM) reconstructions reveal an elongated inchworm-like architecture. Sen1 is composed of an amino terminal helical repeat Sen1 N-terminal (Sen1N) regulatory domain that is flexibly linked to its C-terminal SF1B helicase motor core (Sen1Hel) via an intrinsically disordered tether. In an autoinhibited state, the Sen1Sen1N domain regulates substrate engagement by promoting occlusion of the RNA substrate-binding cleft. The X-ray structure of an activated Sen1Hel engaging single-stranded RNA and ADP-SO4 shows that the enzyme encircles RNA and implicates a single-nucleotide power stroke in the Sen1 RNA translocation mechanism. Together, our data unveil dynamic protein-protein and protein-RNA interfaces underpinning helicase regulation and inactivation of human SETX activity by RNA-binding-deficient mutants in ataxia with oculomotor apraxia 2 neurodegenerative disease.
Keywords: DNA repair; Helicase; R-loop; RNA-DNA hybrid; SETX; SF1B; Sen1; X-ray crystallography; cryo-EM; senataxin; transcription.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Steinmetz EJ, Warren CL, Kuehner JN, Panbehi B, Ansari AZ, and Brow DA (2006). Genome-wide distribution of yeast RNA polymerase II and its control by Sen1 helicase. Mol Cell 24, 735–746. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
