

Characterizing the regulatory Fas (CD95) epitope critical for agonist antibody targeting and CAR-T bystander function in ovarian cancer
- PMID: 37838774
- PMCID: PMC10657439
- DOI: 10.1038/s41418-023-01229-7
Characterizing the regulatory Fas (CD95) epitope critical for agonist antibody targeting and CAR-T bystander function in ovarian cancer
Abstract
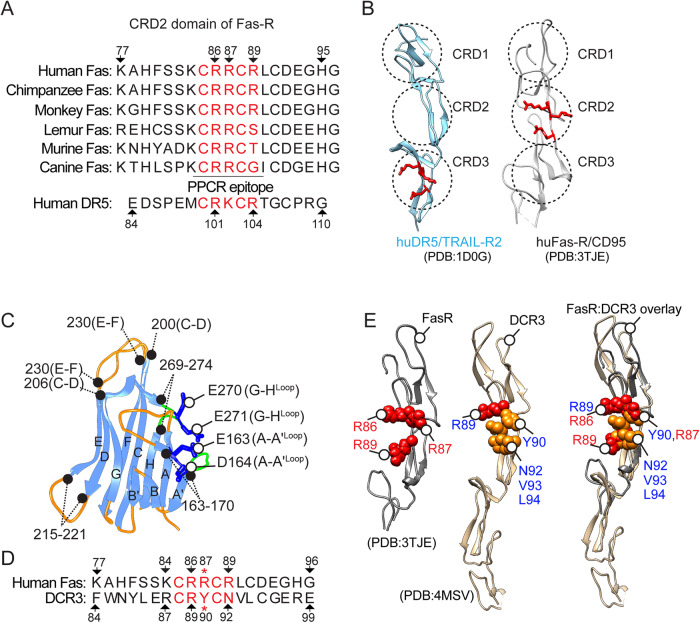
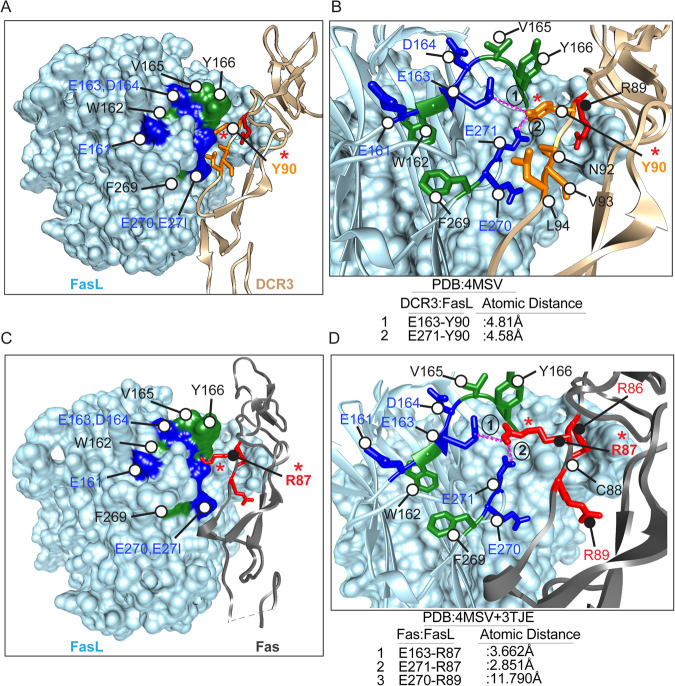
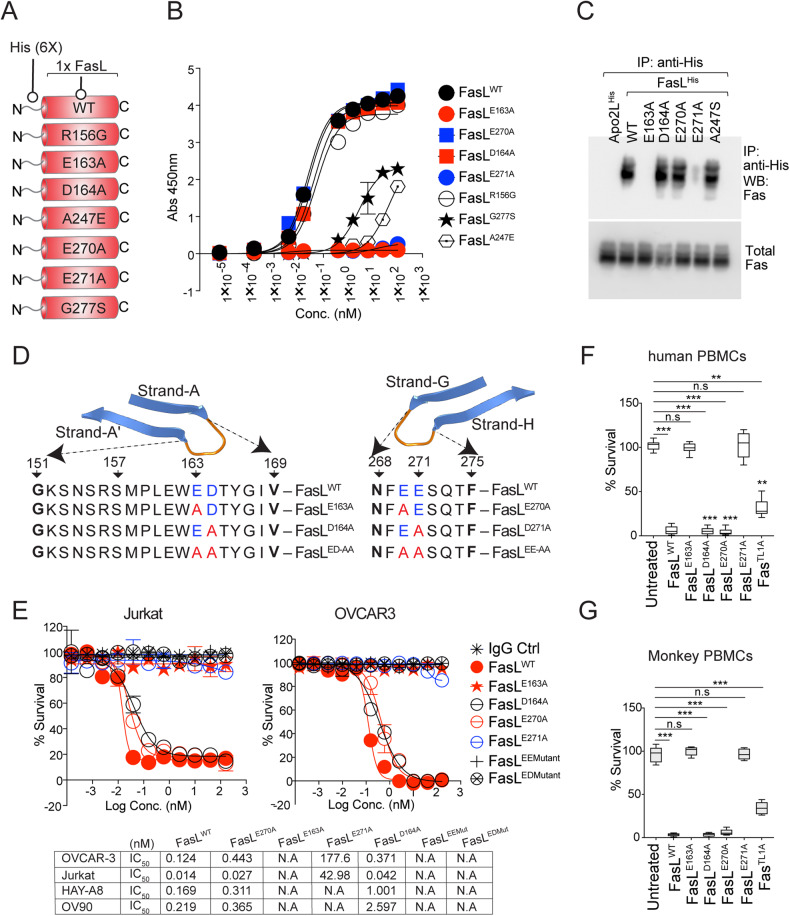
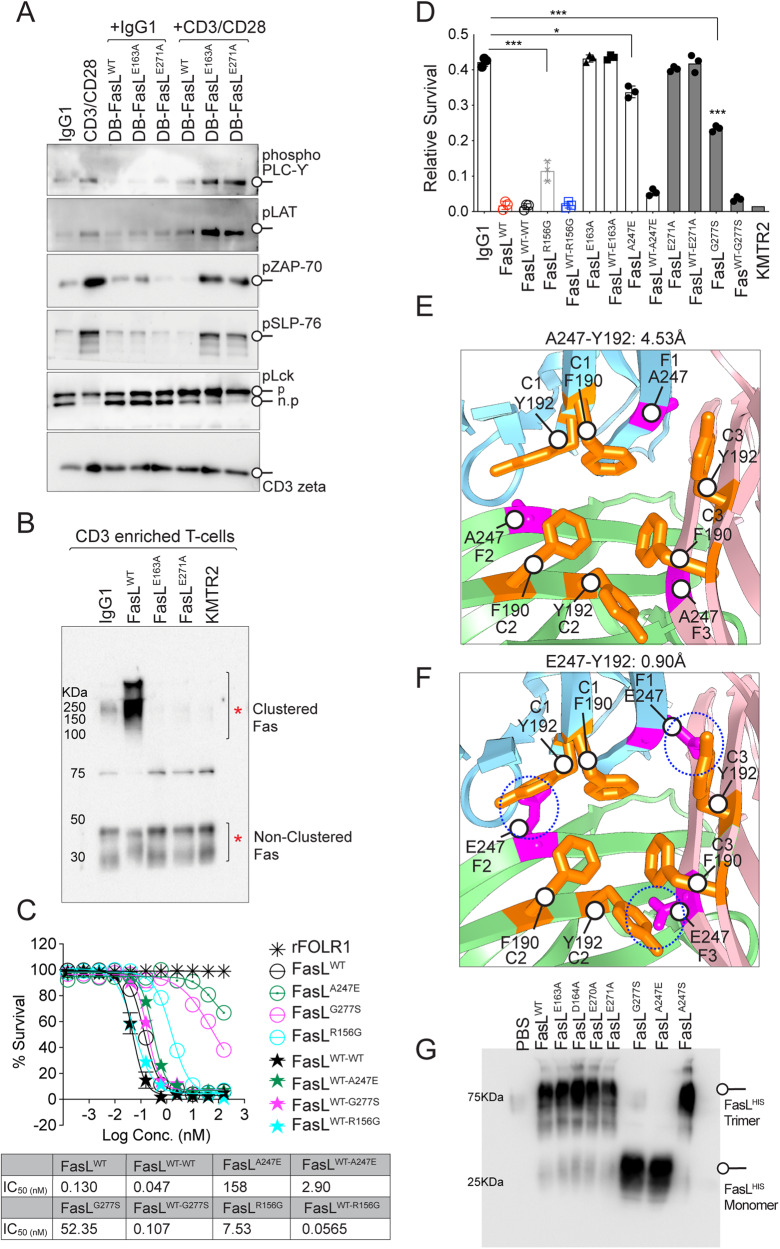
Receptor clustering is the most critical step to activate extrinsic apoptosis by death receptors belonging to the TNF superfamily. Although clinically unsuccessful, using agonist antibodies, the death receptors-5 remains extensively studied from a cancer therapeutics perspective. However, despite its regulatory role and elevated function in ovarian and other solid tumors, another tumor-enriched death receptor called Fas (CD95) remained undervalued in cancer immunotherapy until recently, when its role in off-target tumor killing by CAR-T therapies was imperative. By comprehensively analyzing structure studies in the context of the binding epitope of FasL and various preclinical Fas agonist antibodies, we characterize a highly significant patch of positively charged residue epitope (PPCR) in its cysteine-rich domain 2 of Fas. PPCR engagement is indispensable for superior Fas agonist signaling and CAR-T bystander function in ovarian tumor models. A single-point mutation in FasL or Fas that interferes with the PPCR engagement inhibited apoptotic signaling in tumor cells and T cells. Furthermore, considering that clinical and immunological features of the autoimmune lymphoproliferative syndrome (ALPS) are directly attributed to homozygous mutations in FasL, we reveal differential mechanistic details of FasL/Fas clustering at the PPCR interface compared to described ALPS mutations. As Fas-mediated bystander killing remains vital to the success of CAR-T therapies in tumors, our findings highlight the therapeutic analytical design for potentially effective Fas-targeting strategies using death agonism to improve cancer immunotherapy in ovarian and other solid tumors.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
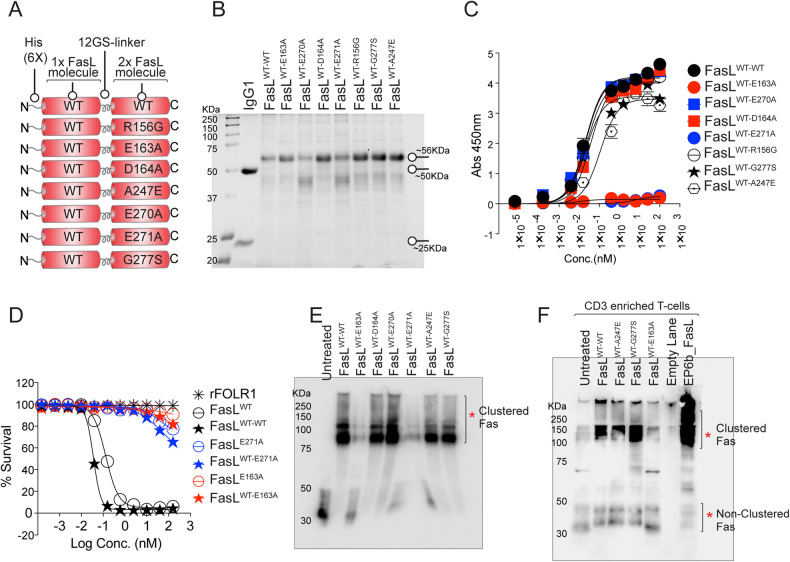
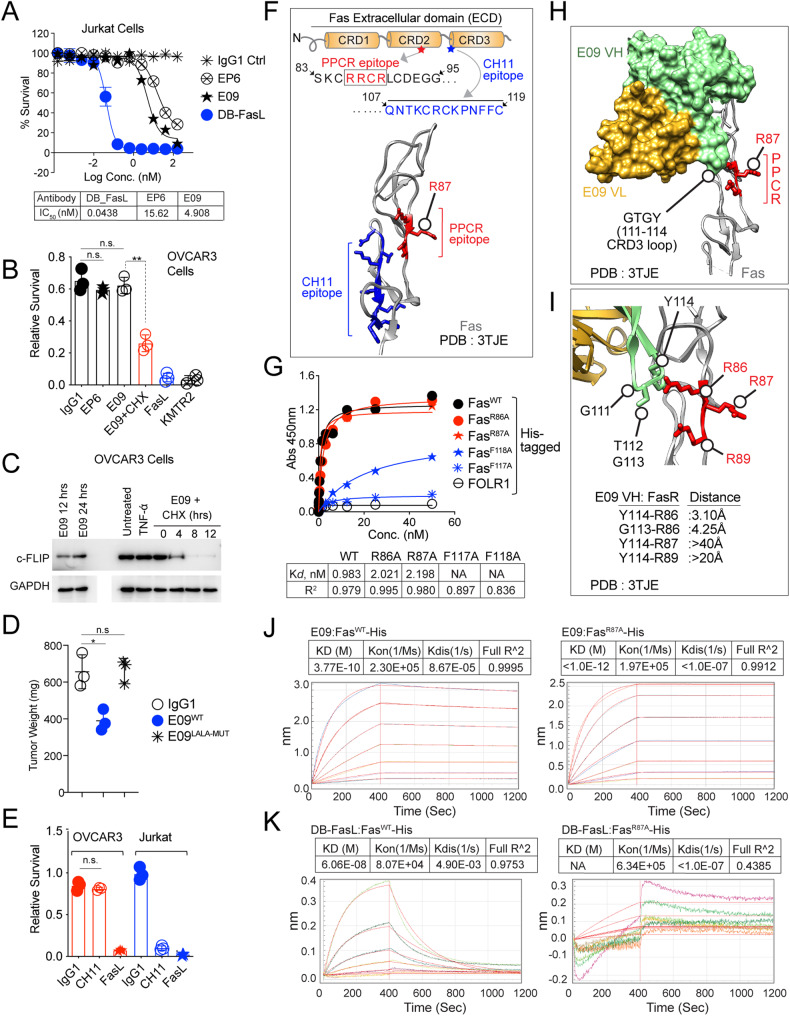
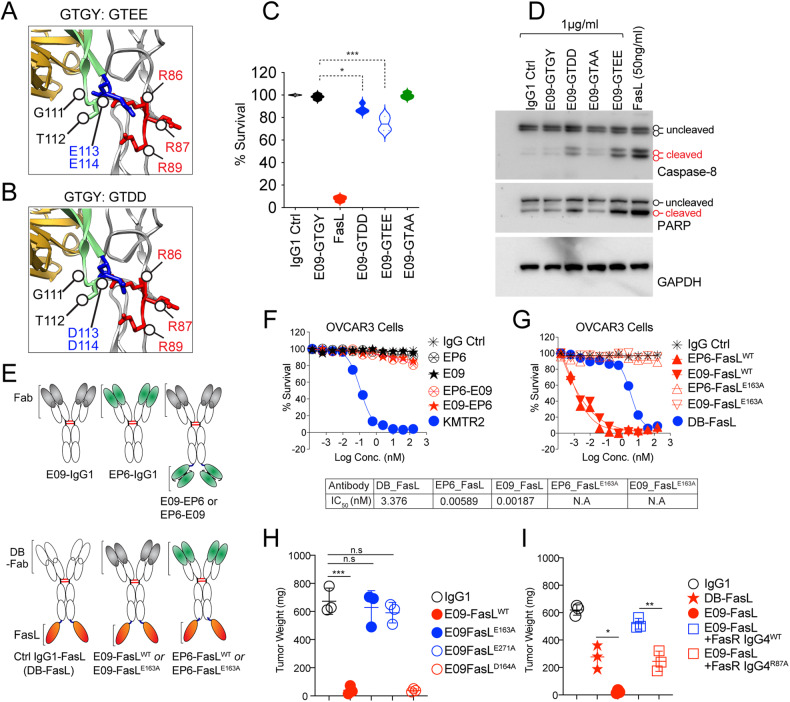
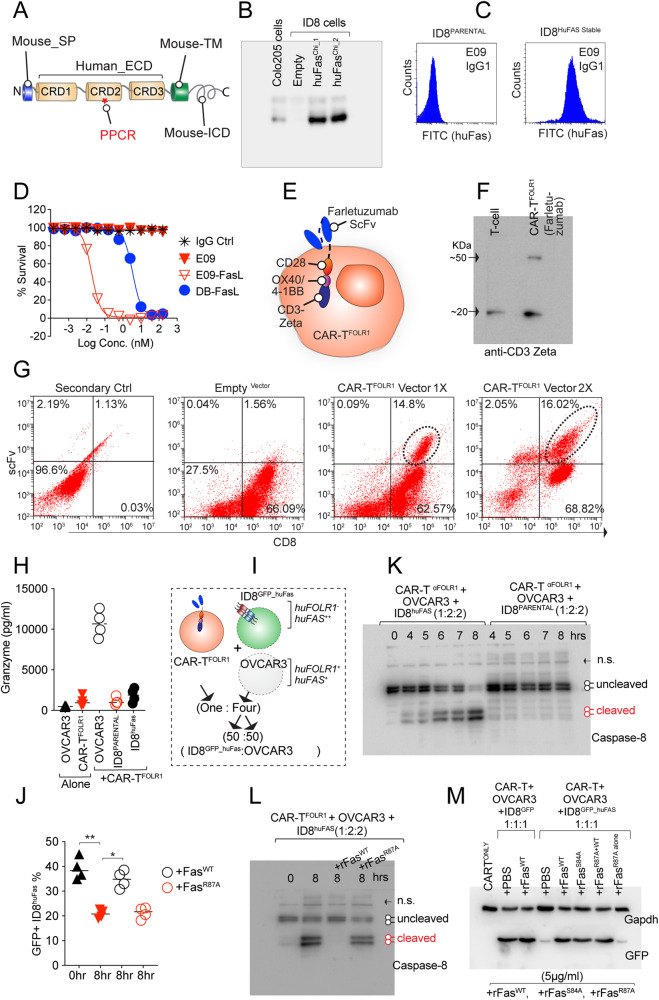
References
-
- Ashkenazi A. Directing cancer cells to self-destruct with pro-apoptotic receptor agonists. Nat Rev Drug Discov. 2008;7:1001–12. - PubMed
-
- Adams C, Totpal K, Lawrence D, Marsters S, Pitti R, Yee S, et al. Structural and functional analysis of the interaction between the agonistic monoclonal antibody Apomab and the proapoptotic receptor DR5. Cell Death Differ. 2008;15:751–61. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
