

Cyanobacteriochromes from Gloeobacterales Provide New Insight into the Diversification of Cyanobacterial Photoreceptors
- PMID: 37839679
- PMCID: PMC11218821
- DOI: 10.1016/j.jmb.2023.168313
Cyanobacteriochromes from Gloeobacterales Provide New Insight into the Diversification of Cyanobacterial Photoreceptors
Abstract
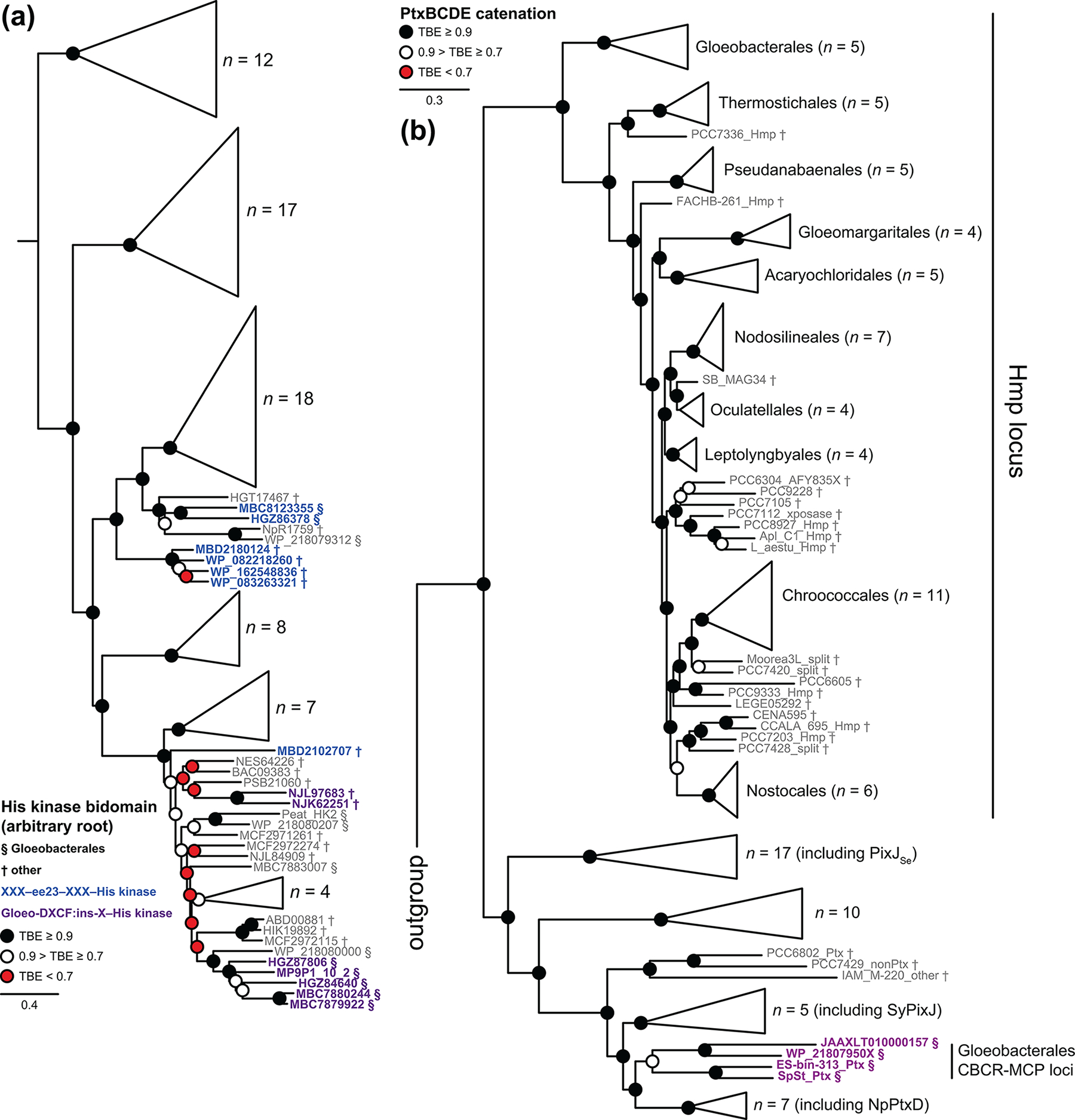
The phytochrome superfamily comprises three groups of photoreceptors sharing a conserved GAF (cGMP-specific phosphodiesterases, cyanobacterial adenylate cyclases, and formate hydrogen lyase transcription activator FhlA) domain that uses a covalently attached linear tetrapyrrole (bilin) chromophore to sense light. Knotted red/far-red phytochromes are widespread in both bacteria and eukaryotes, but cyanobacteria also contain knotless red/far-red phytochromes and cyanobacteriochromes (CBCRs). Unlike typical phytochromes, CBCRs require only the GAF domain for bilin binding, chromophore ligation, and full, reversible photoconversion. CBCRs can sense a wide range of wavelengths (ca. 330-750 nm) and can regulate phototaxis, second messenger metabolism, and optimization of the cyanobacterial light-harvesting apparatus. However, the origins of CBCRs are not well understood: we do not know when or why CBCRs evolved, or what selective advantages led to retention of early CBCRs in cyanobacterial genomes. In the current work, we use the increasing availability of genomes and metagenome-assembled-genomes from early-branching cyanobacteria to explore the origins of CBCRs. We reaffirm the earliest branches in CBCR evolution. We also show that early-branching cyanobacteria contain late-branching CBCRs, implicating early appearance of CBCRs during cyanobacterial evolution. Moreover, we show that early-branching CBCRs behave as integrators of light and pH, providing a potential unique function for early CBCRs that led to their retention and subsequent diversification. Our results thus provide new insight into the origins of these diverse cyanobacterial photoreceptors.
Keywords: chromatic acclimation; photosynthesis; phototaxis; phycocyanobilin; phytochrome.
Copyright © 2023 The Author(s). Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures










Similar articles
-
Cyanobacteriochromes: A Rainbow of Photoreceptors.Annu Rev Microbiol. 2024 Nov;78(1):61-81. doi: 10.1146/annurev-micro-041522-094613. Epub 2024 Nov 7. Annu Rev Microbiol. 2024. PMID: 38848579 Free PMC article. Review.
-
Teal-light absorbing cyanobacterial phytochrome superfamily provides insights into the diverse mechanisms of spectral tuning and facilitates the engineering of photoreceptors for optogenetic tools.Int J Biol Macromol. 2024 Aug;274(Pt 2):133407. doi: 10.1016/j.ijbiomac.2024.133407. Epub 2024 Jun 24. Int J Biol Macromol. 2024. PMID: 38925190
-
Cyanobacteriochrome Photoreceptors Lacking the Canonical Cys Residue.Biochemistry. 2016 Dec 20;55(50):6981-6995. doi: 10.1021/acs.biochem.6b00940. Epub 2016 Dec 9. Biochemistry. 2016. PMID: 27935696
-
Diverse two-cysteine photocycles in phytochromes and cyanobacteriochromes.Proc Natl Acad Sci U S A. 2011 Jul 19;108(29):11854-9. doi: 10.1073/pnas.1107844108. Epub 2011 Jun 28. Proc Natl Acad Sci U S A. 2011. PMID: 21712441 Free PMC article.
-
Cyanobacteriochromes: a new superfamily of tetrapyrrole-binding photoreceptors in cyanobacteria.Photochem Photobiol Sci. 2008 Oct;7(10):1159-67. doi: 10.1039/b802660m. Epub 2008 Aug 18. Photochem Photobiol Sci. 2008. PMID: 18846279 Review.
Cited by
-
Cyanobacteriochromes: A Rainbow of Photoreceptors.Annu Rev Microbiol. 2024 Nov;78(1):61-81. doi: 10.1146/annurev-micro-041522-094613. Epub 2024 Nov 7. Annu Rev Microbiol. 2024. PMID: 38848579 Free PMC article. Review.
-
Red/green cyanobacteriochromes acquire isomerization from phycocyanobilin to phycoviolobilin.Protein Sci. 2024 Aug;33(8):e5132. doi: 10.1002/pro.5132. Protein Sci. 2024. PMID: 39072823 Free PMC article.
-
Dual-Cys bacteriophytochromes: intermediates in cyanobacterial phytochrome evolution?FEBS J. 2025 Mar;292(5):1197-1216. doi: 10.1111/febs.17395. Epub 2025 Jan 13. FEBS J. 2025. PMID: 39801362 Free PMC article.
-
Circular dichroism spectroscopy reveals multiple phytochrome photoproducts in equilibrium.Photochem Photobiol Sci. 2025 Aug;24(8):1351-1371. doi: 10.1007/s43630-025-00763-2. Epub 2025 Jul 18. Photochem Photobiol Sci. 2025. PMID: 40681765 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
