

Sex differences in vocal learning ability in songbirds are linked with differences in flexible rhythm pattern perception
- PMID: 37842009
- PMCID: PMC10569135
- DOI: 10.1016/j.anbehav.2023.05.001
Sex differences in vocal learning ability in songbirds are linked with differences in flexible rhythm pattern perception
Abstract
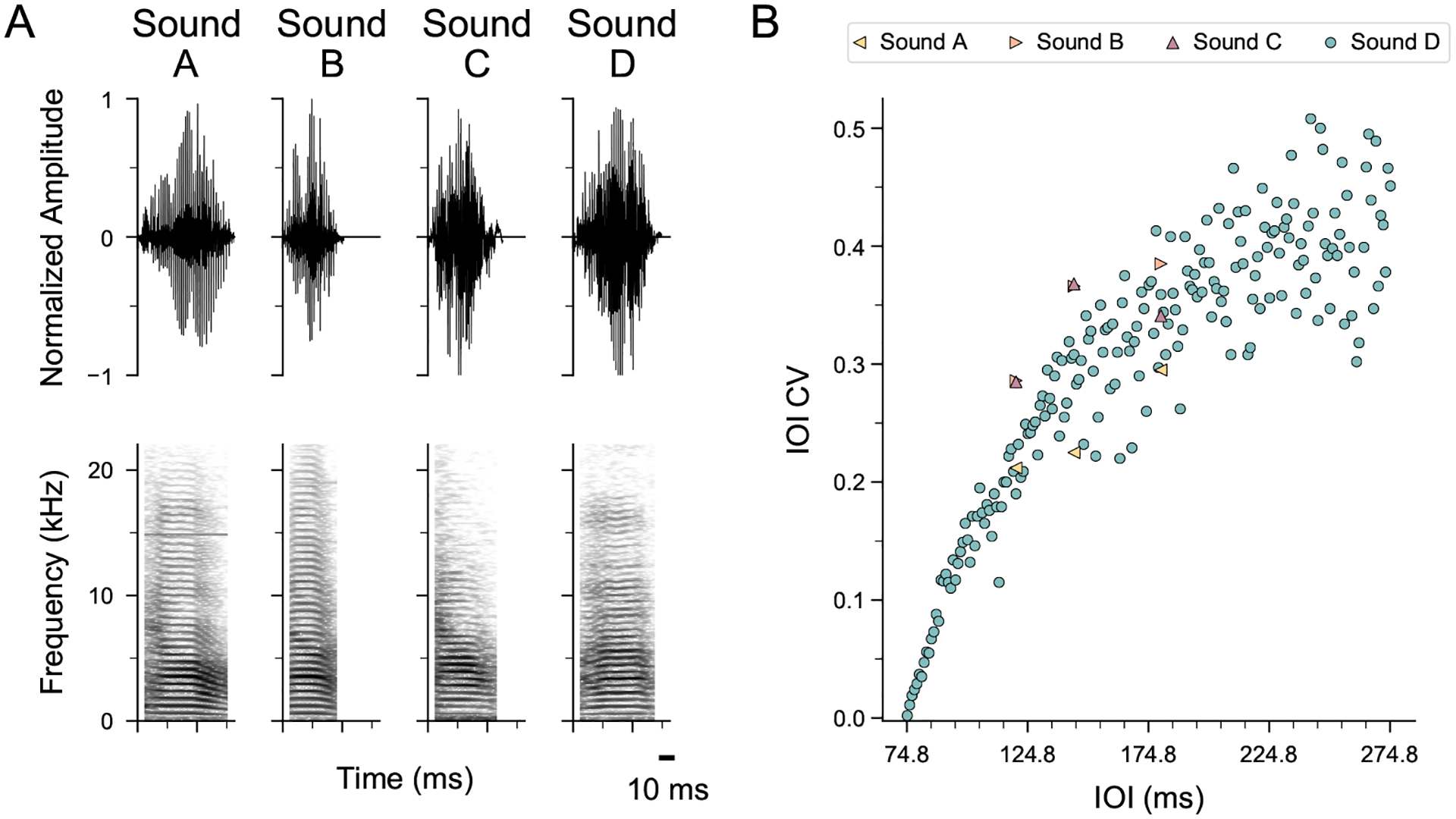
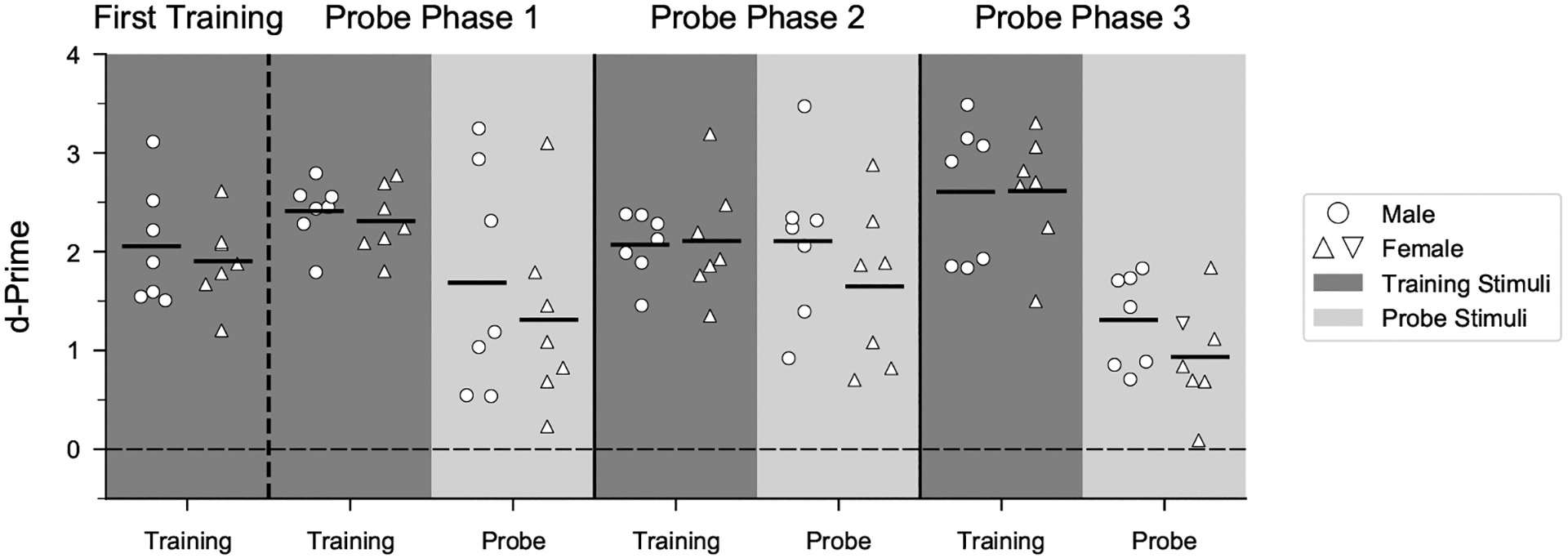
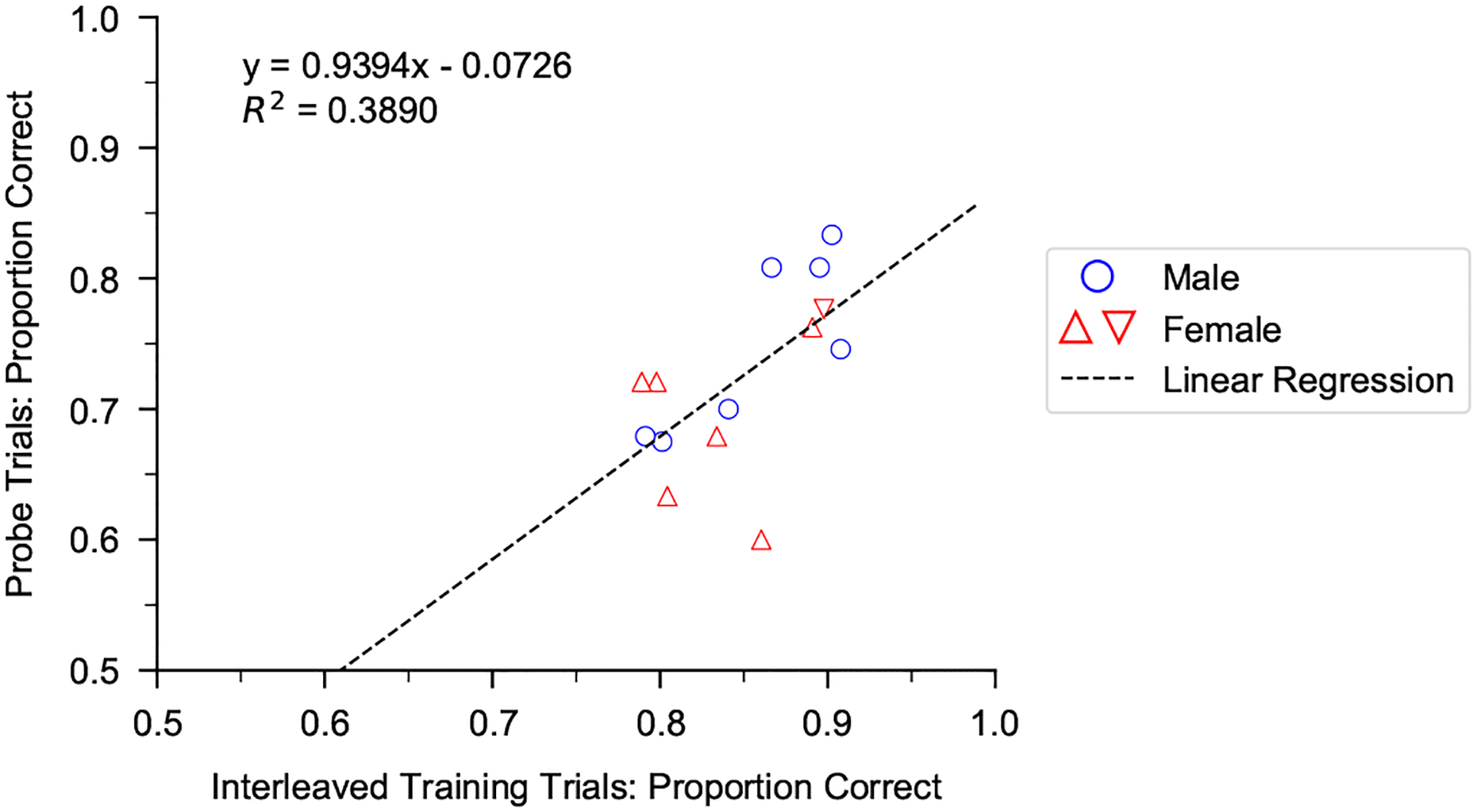
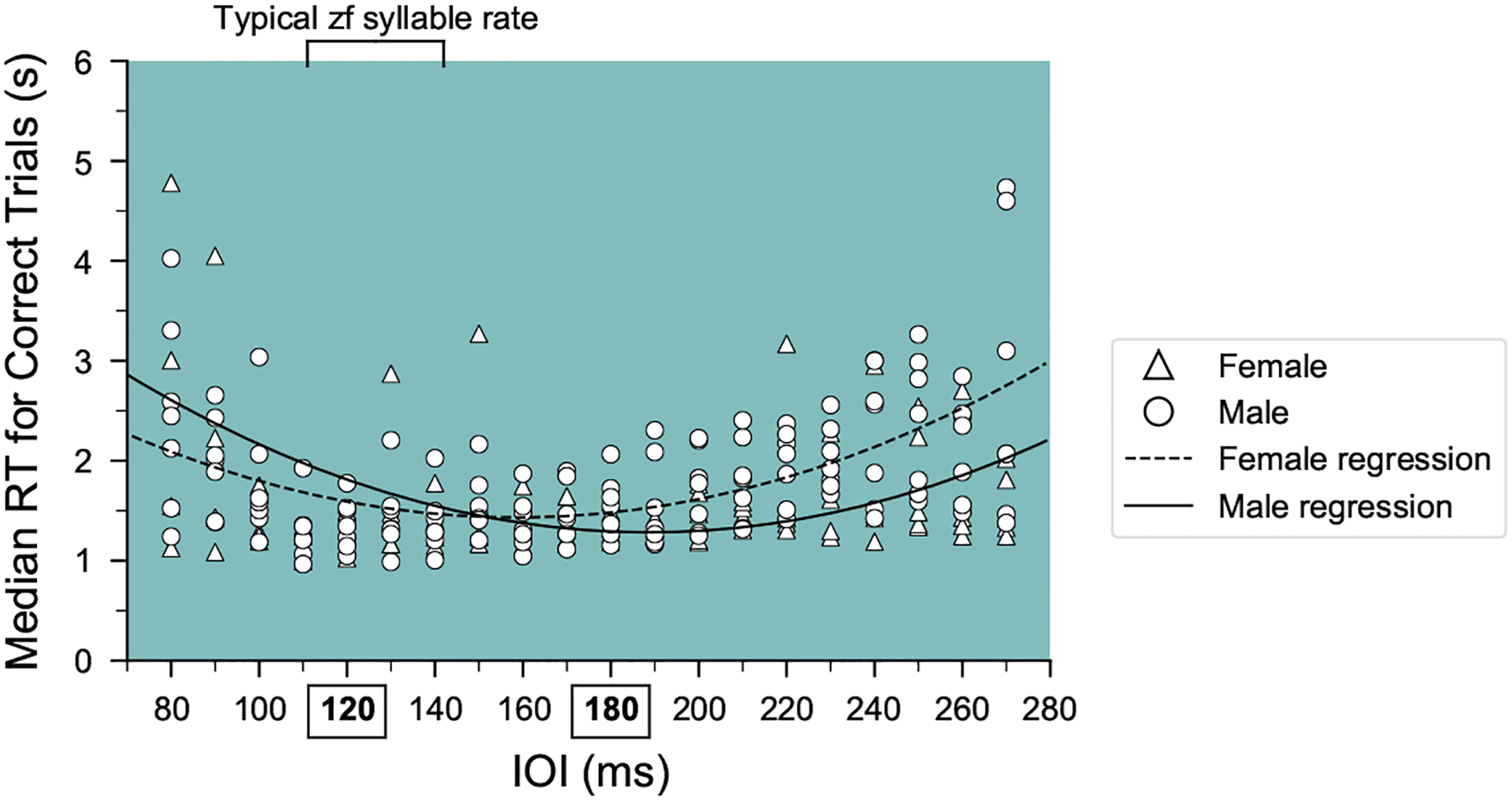
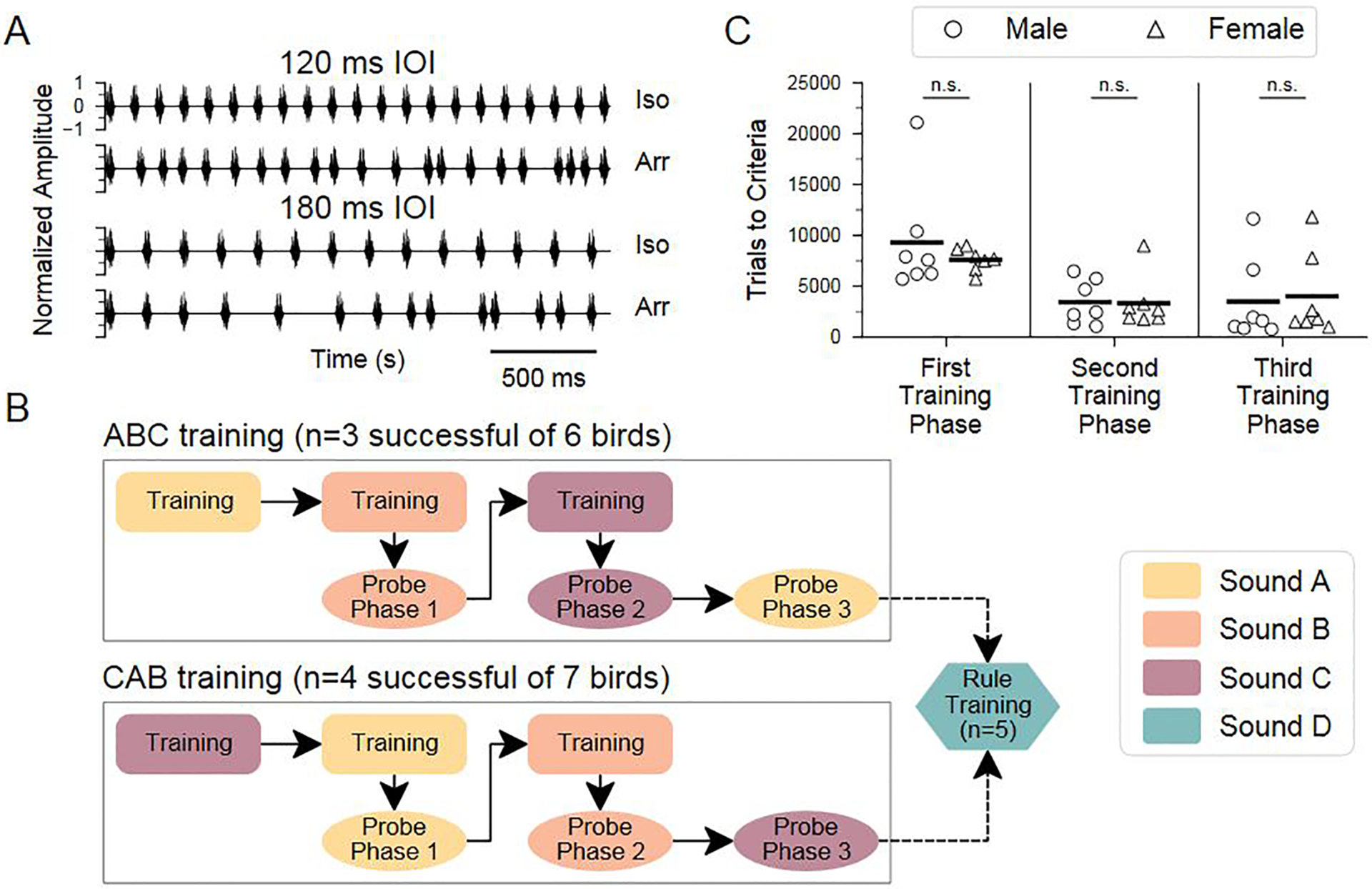
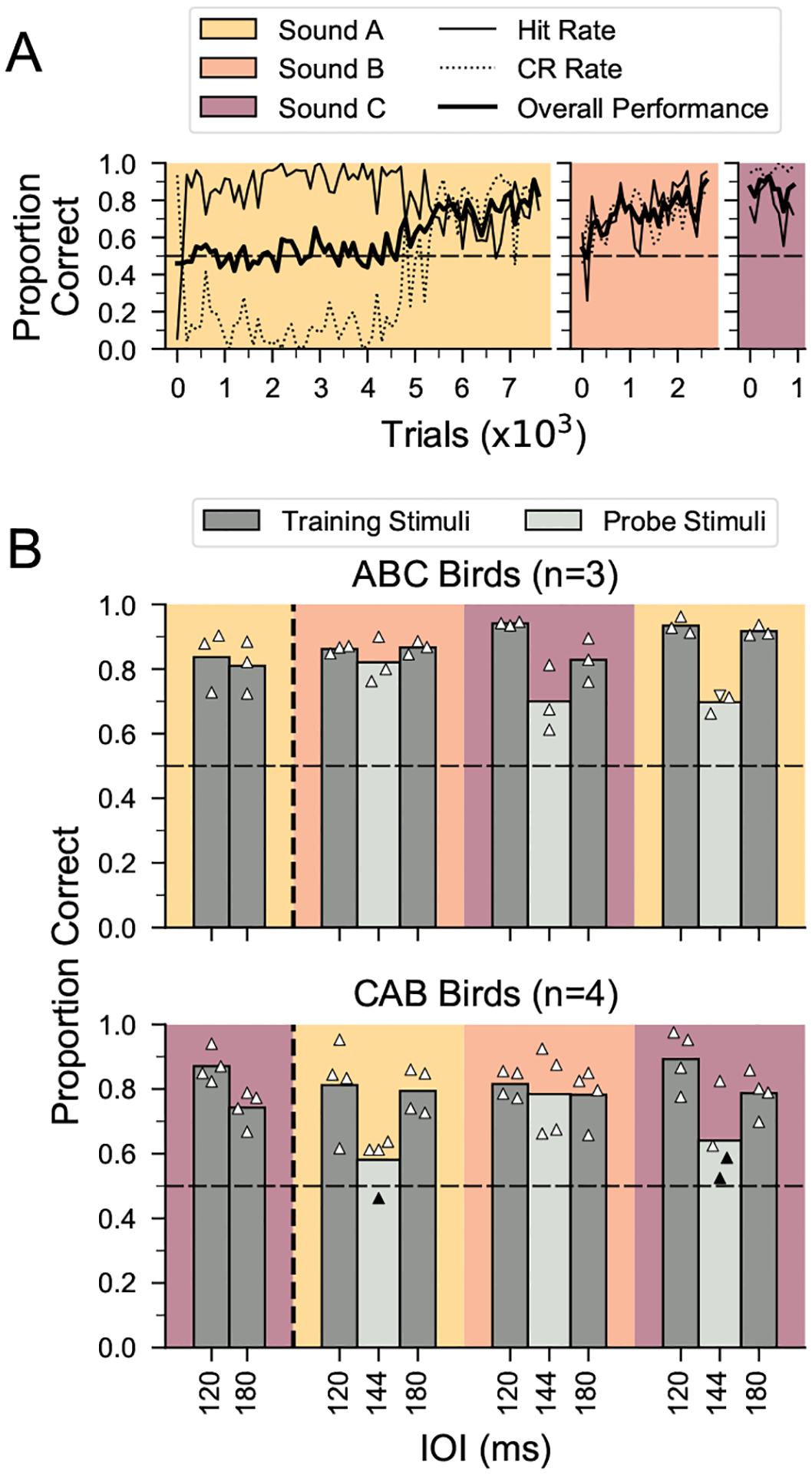
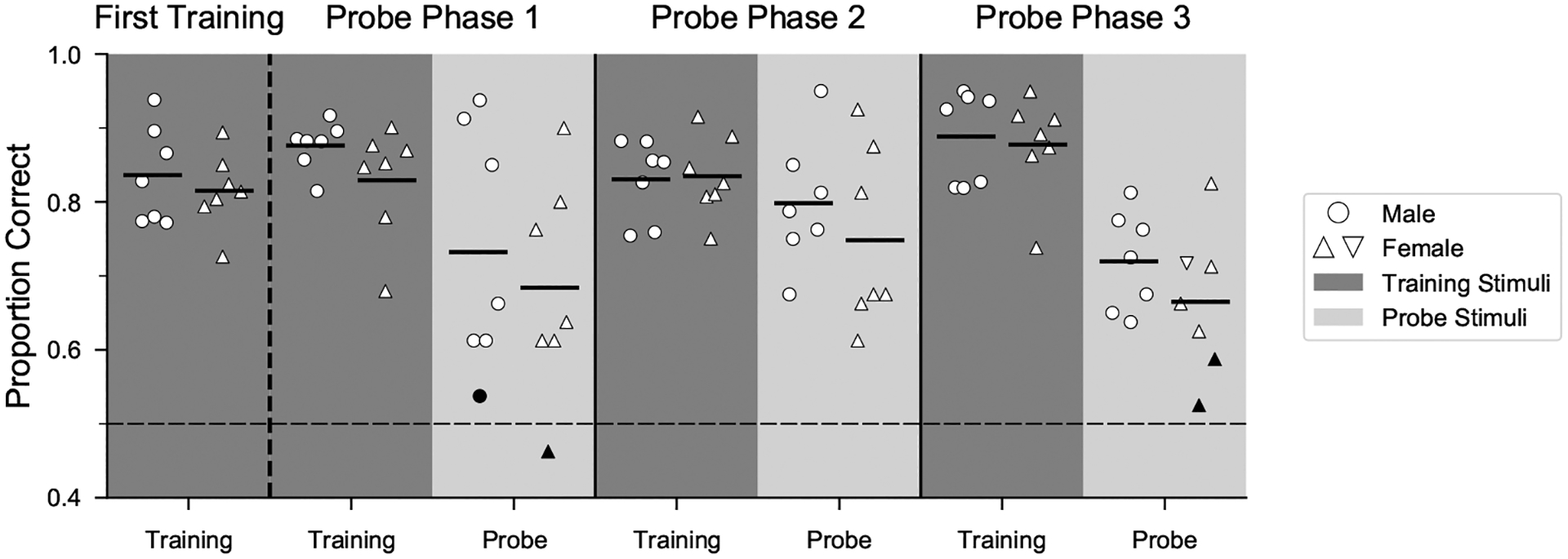
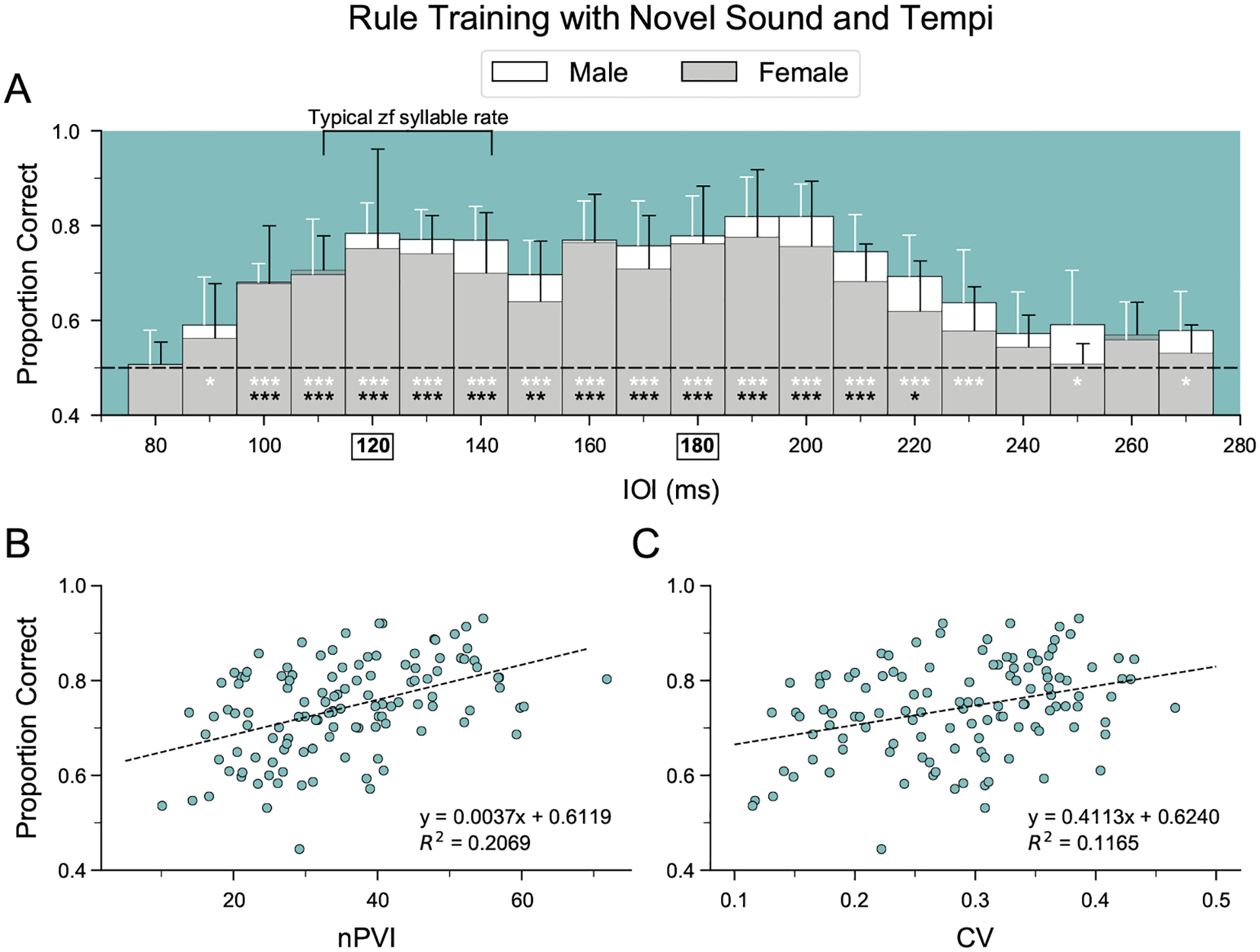
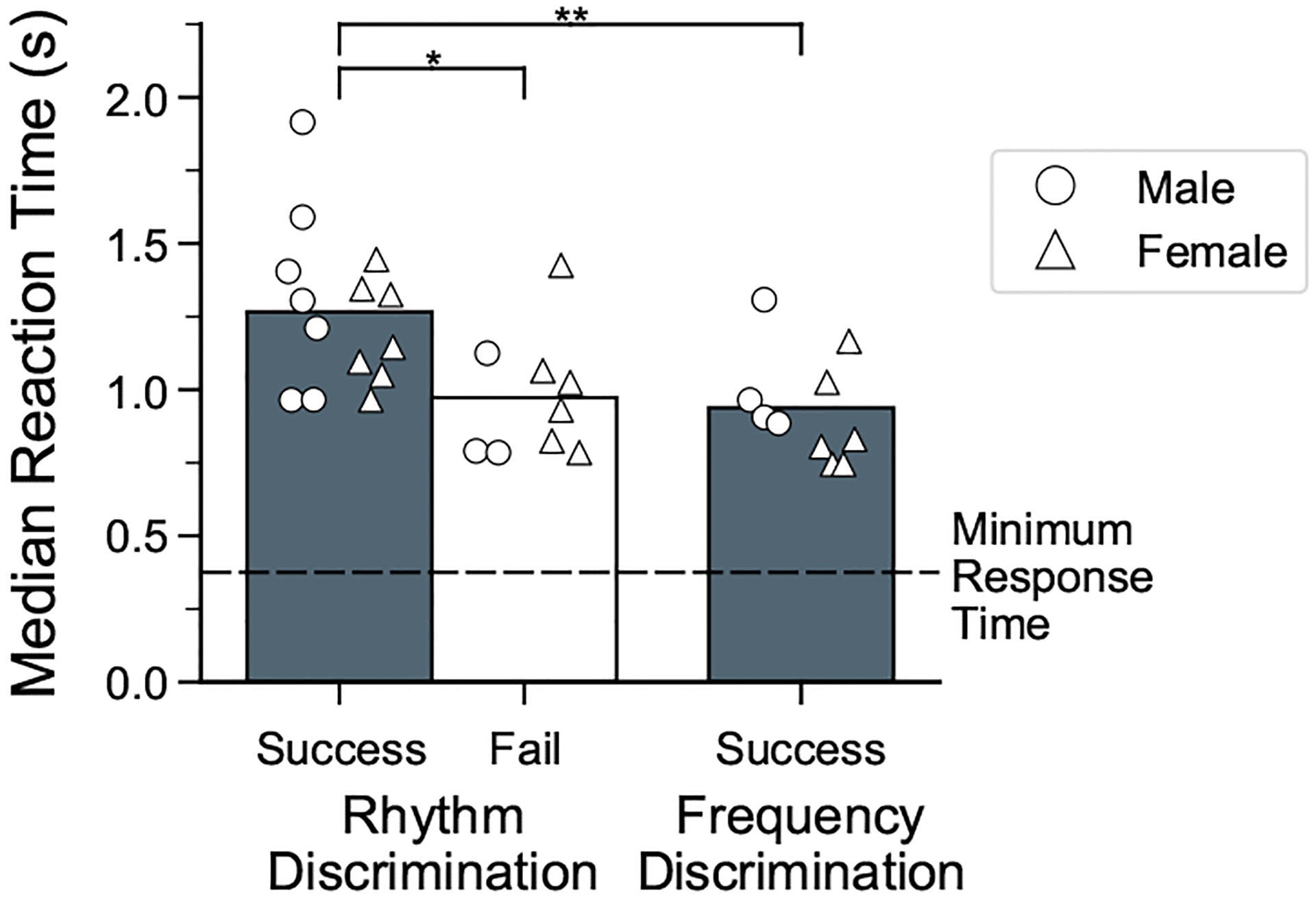
Humans readily recognize familiar rhythmic patterns, such as isochrony (equal timing between events) across a wide range of rates. This reflects a facility with perceiving the relative timing of events, not just absolute interval durations. Several lines of evidence suggest this ability is supported by precise temporal predictions arising from forebrain auditory-motor interactions. We have shown previously that male zebra finches, Taeniopygia guttata, which possess specialized auditory-motor networks and communicate with rhythmically patterned sequences, share our ability to flexibly recognize isochrony across rates. To test the hypothesis that flexible rhythm pattern perception is linked to vocal learning, we ask whether female zebra finches, which do not learn to sing, can also recognize global temporal patterns. We find that females can flexibly recognize isochrony across a wide range of rates but perform slightly worse than males on average. These findings are consistent with recent work showing that while females have reduced forebrain song regions, the overall network connectivity of vocal premotor regions is similar to males and may support predictions of upcoming events. Comparative studies of male and female songbirds thus offer an opportunity to study how individual differences in auditory-motor connectivity influence perception of relative timing, a hallmark of human music perception.
Keywords: auditory perception; auditory–motor interaction; comparative cognition; isochrony; sensorimotor circuit; zebra finch.
Conflict of interest statement
Declaration of Interest None.
Figures

Similar articles
-
Vocal learning and flexible rhythm pattern perception are linked: Evidence from songbirds.Proc Natl Acad Sci U S A. 2021 Jul 20;118(29):e2026130118. doi: 10.1073/pnas.2026130118. Epub 2021 Jul 16. Proc Natl Acad Sci U S A. 2021. PMID: 34272278 Free PMC article.
-
Finding the Beat: From Socially Coordinated Vocalizations in Songbirds to Rhythmic Entrainment in Humans.Front Hum Neurosci. 2016 Jun 6;10:255. doi: 10.3389/fnhum.2016.00255. eCollection 2016. Front Hum Neurosci. 2016. PMID: 27375455 Free PMC article. Review.
-
Female zebra finches do not sing yet share neural pathways necessary for singing in males.J Comp Neurol. 2019 Mar 1;527(4):843-855. doi: 10.1002/cne.24569. Epub 2018 Dec 4. J Comp Neurol. 2019. PMID: 30370534
-
Individual vocal recognition in zebra finches relies on song syllable structure rather than song syllable order.J Exp Biol. 2020 May 7;223(Pt 9):jeb220087. doi: 10.1242/jeb.220087. J Exp Biol. 2020. PMID: 32253282
-
The zebra finch, Taeniopygia guttata: an avian model for investigating the neurobiological basis of vocal learning.Cold Spring Harb Protoc. 2014 Oct 23;2014(12):1237-42. doi: 10.1101/pdb.emo084574. Cold Spring Harb Protoc. 2014. PMID: 25342070 Free PMC article. Review.
Cited by
-
Rhythmic coordination in rapid duets of little crakes: cooperative signalling without shared experience.Proc Biol Sci. 2025 Jul;292(2051):20251100. doi: 10.1098/rspb.2025.1100. Epub 2025 Jul 23. Proc Biol Sci. 2025. PMID: 40695354 Free PMC article.
-
Beat-based dancing to music has evolutionary foundations in advanced vocal learning.BMC Neurosci. 2024 Nov 6;25(1):65. doi: 10.1186/s12868-024-00843-6. BMC Neurosci. 2024. PMID: 39506663 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources