

Prdm16 mutation determines sex-specific cardiac metabolism and identifies two novel cardiac metabolic regulators
- PMID: 37842925
- PMCID: PMC10874277
- DOI: 10.1093/cvr/cvad154
Prdm16 mutation determines sex-specific cardiac metabolism and identifies two novel cardiac metabolic regulators
Abstract
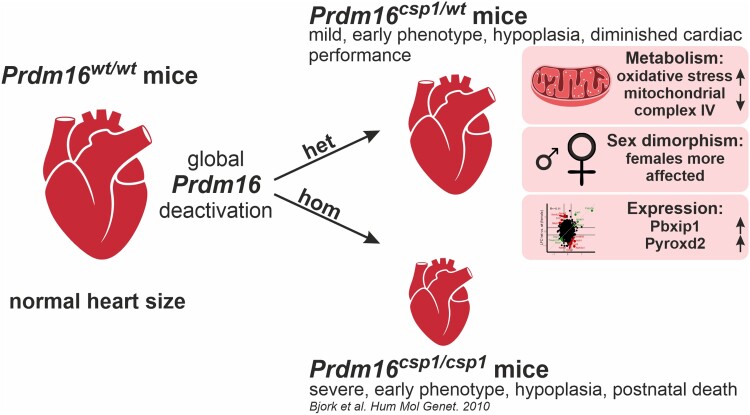
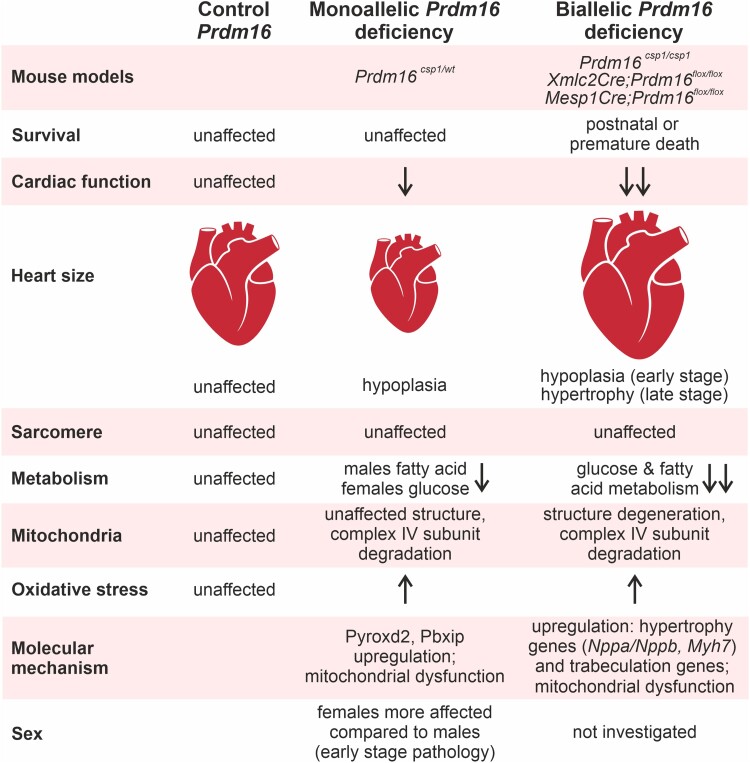
Aims: Mutation of the PRDM16 gene causes human dilated and non-compaction cardiomyopathy. The PRDM16 protein is a transcriptional regulator that affects cardiac development via Tbx5 and Hand1, thus regulating myocardial structure. The biallelic inactivation of Prdm16 induces severe cardiac dysfunction with post-natal lethality and hypertrophy in mice. The early pathological events that occur upon Prdm16 inactivation have not been explored.
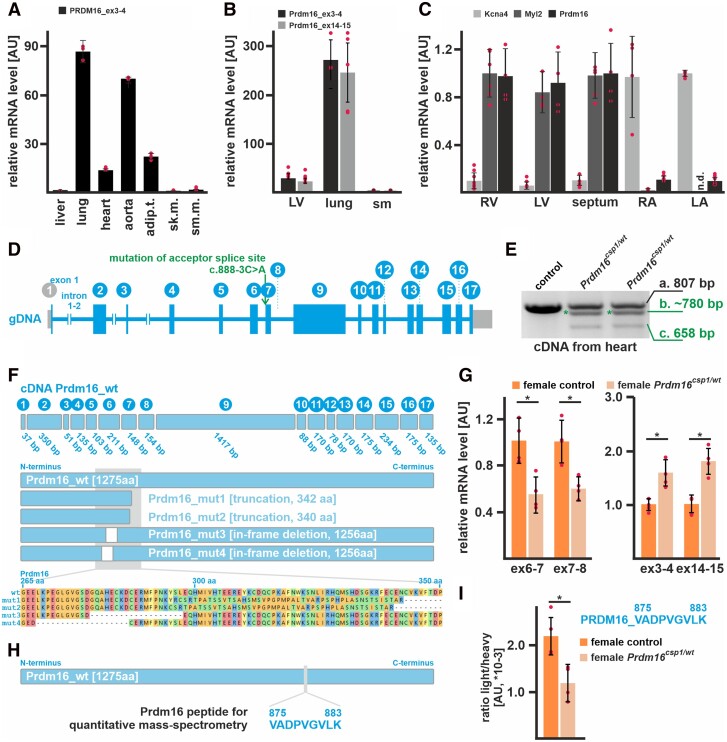
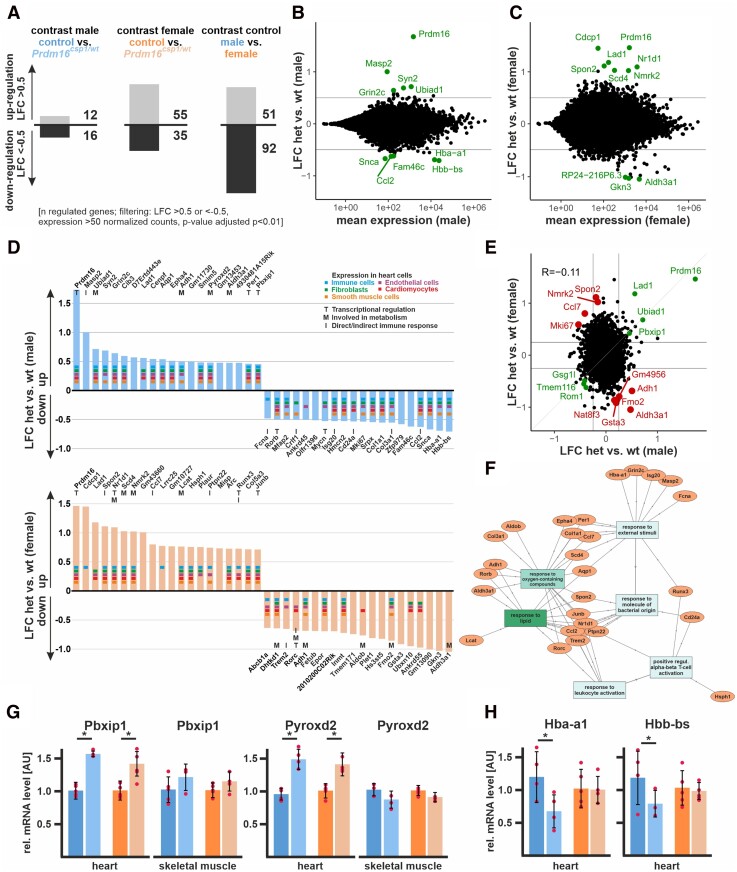
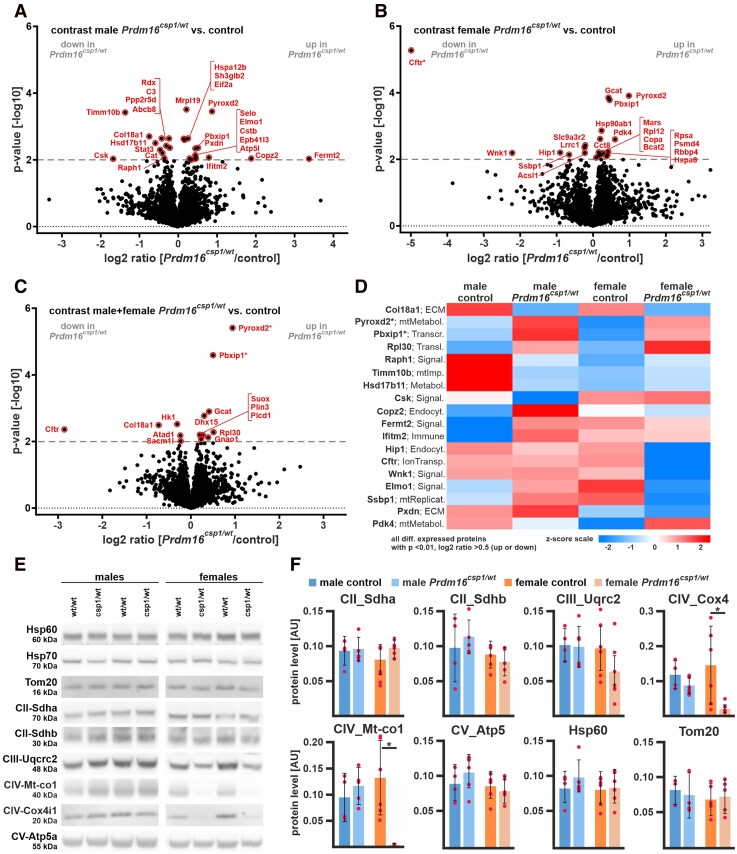
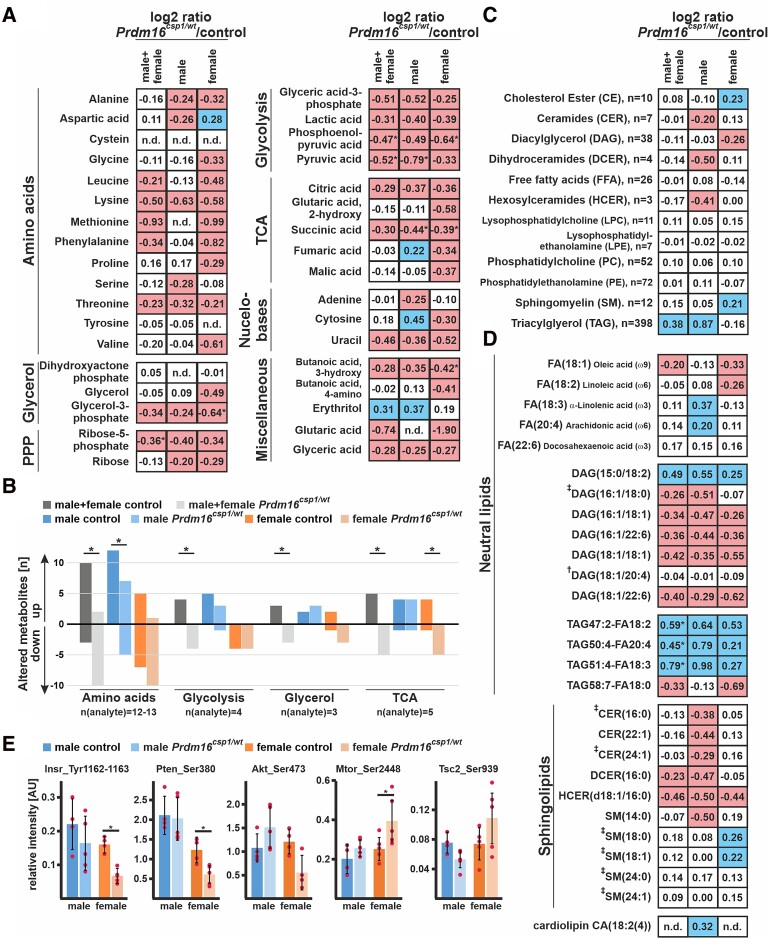
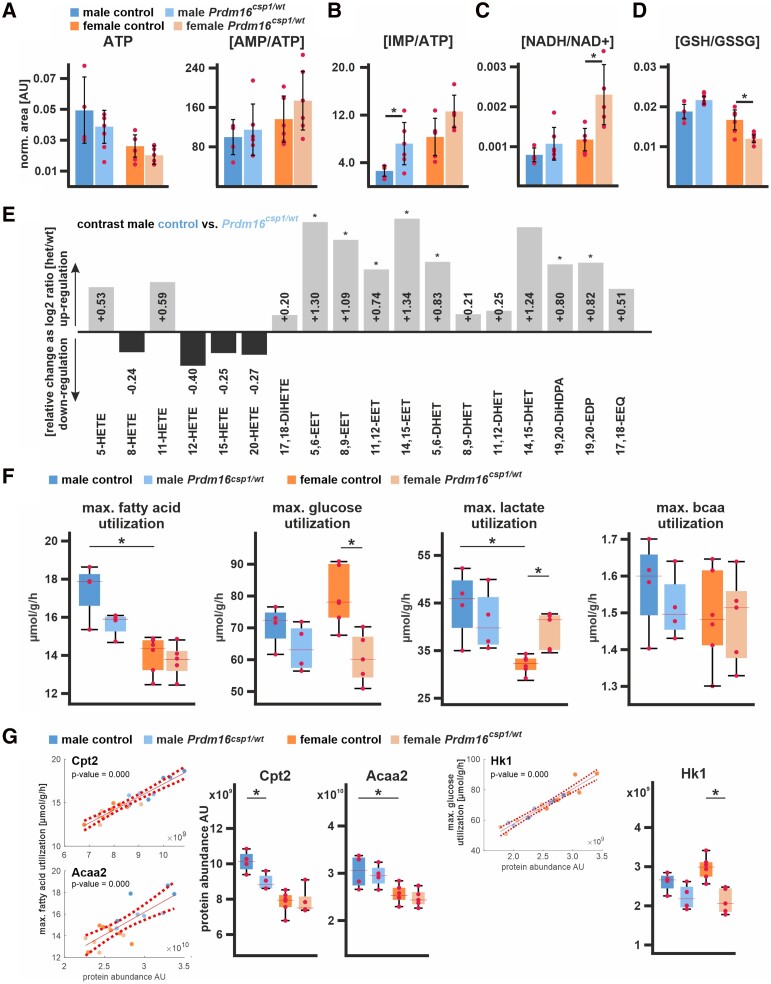
Methods and results: This study performed in-depth pathophysiological and molecular analyses of male and female Prdm16csp1/wt mice that carry systemic, monoallelic Prdm16 gene inactivation. We systematically assessed early molecular changes through transcriptomics, proteomics, and metabolomics. Kinetic modelling of cardiac metabolism was performed in silico with CARDIOKIN. Prdm16csp1/wt mice are viable up to 8 months, develop hypoplastic hearts, and diminished systolic performance that is more pronounced in female mice. Prdm16csp1/wt cardiac tissue of both sexes showed reductions in metabolites associated with amino acid as well as glycerol metabolism, glycolysis, and the tricarboxylic acid cycle. Prdm16csp1/wt cardiac tissue revealed diminished glutathione (GSH) and increased inosine monophosphate (IMP) levels indicating oxidative stress and a dysregulated energetics, respectively. An accumulation of triacylglycerides exclusively in male Prdm16csp1/wt hearts suggests a sex-specific metabolic adaptation. Metabolic modelling using CARDIOKIN identified a reduction in fatty acid utilization in males as well as lower glucose utilization in female Prdm16csp1/wt cardiac tissue. On the level of transcripts and protein expression, Prdm16csp1/wt hearts demonstrate an up-regulation of pyridine nucleotide-disulphide oxidoreductase domain 2 (Pyroxd2) and the transcriptional regulator pre-B-cell leukaemia transcription factor interacting protein 1 (Pbxip1). The strongest concordant transcriptional up-regulation was detected for Prdm16 itself, probably through an autoregulatory mechanism.
Conclusions: Monoallelic, global Prdm16 mutation diminishes cardiac performance in Prdm16csp1/wt mice. Metabolic alterations and transcriptional dysregulation in Prdm16csp1/wt affect cardiac tissue. Female Prdm16csp1/wt mice develop a more pronounced phenotype, indicating sexual dimorphism at this early pathological window. This study suggests that metabolic dysregulation is an early event in the PRDM16 associated cardiac pathology.
Keywords: Cardiomyopathy; Metabolism; Mutation; Prdm16.
© The Author(s) 2023. Published by Oxford University Press on behalf of the European Society of Cardiology.
Conflict of interest statement
Conflict of interest: All authors declare to have no conflict of interest related to this manuscript. The patent application EP21174633 with the title ‘Computer assisted method for the evaluation of cardiac metabolism’ was filed by Charité—Universitätsmedizin Berlin as the employer of N.B. and Titus Kuehne, with both holding inventorship for this patent application.
Figures

Similar articles
-
Cardiac-specific inactivation of Prdm16 effects cardiac conduction abnormalities and cardiomyopathy-associated phenotypes.Am J Physiol Heart Circ Physiol. 2020 Apr 1;318(4):H764-H777. doi: 10.1152/ajpheart.00647.2019. Epub 2020 Feb 21. Am J Physiol Heart Circ Physiol. 2020. PMID: 32083975
-
PRDM16 Is a Compact Myocardium-Enriched Transcription Factor Required to Maintain Compact Myocardial Cardiomyocyte Identity in Left Ventricle.Circulation. 2022 Feb 22;145(8):586-602. doi: 10.1161/CIRCULATIONAHA.121.056666. Epub 2021 Dec 17. Circulation. 2022. PMID: 34915728 Free PMC article.
-
A cardiac-null mutation of Prdm16 causes hypotension in mice with cardiac hypertrophy via increased nitric oxide synthase 1.PLoS One. 2022 Jul 21;17(7):e0267938. doi: 10.1371/journal.pone.0267938. eCollection 2022. PLoS One. 2022. PMID: 35862303 Free PMC article.
-
PRDM16 Deletion Is Associated With Sex-dependent Cardiomyopathy and Cardiac Mortality: A Translational, Multi-Institutional Cohort Study.Circ Genom Precis Med. 2023 Aug;16(4):390-400. doi: 10.1161/CIRCGEN.122.003912. Epub 2023 Jul 3. Circ Genom Precis Med. 2023. PMID: 37395136 Free PMC article.
-
The Multifaceted Roles of PRDM16: Adipose Biology and Beyond.Trends Endocrinol Metab. 2016 Jan;27(1):11-23. doi: 10.1016/j.tem.2015.11.005. Epub 2015 Dec 11. Trends Endocrinol Metab. 2016. PMID: 26688472 Review.
Cited by
-
Sex-dependent Pathophysiology and Therapeutic Considerations in Right Heart Disease.Can J Cardiol. 2025 Jun;41(6):1038-1053. doi: 10.1016/j.cjca.2025.02.034. Epub 2025 Mar 5. Can J Cardiol. 2025. PMID: 40054579 Review.
-
PRDM16 determines specification of ventricular cardiomyocytes by suppressing alternative cell fates.Life Sci Alliance. 2024 Sep 20;7(12):e202402719. doi: 10.26508/lsa.202402719. Print 2024 Dec. Life Sci Alliance. 2024. PMID: 39304345 Free PMC article.
-
PRDM16, a new kid on the block in cardiovascular health and disease.Cardiovasc Res. 2025 Jul 31;121(8):1156-1172. doi: 10.1093/cvr/cvaf089. Cardiovasc Res. 2025. PMID: 40439125 Free PMC article. Review.
-
Overexpression of PRDM16 attenuates acute kidney injury progression: genetic and pharmacological approaches.MedComm (2020). 2024 Sep 21;5(10):e737. doi: 10.1002/mco2.737. eCollection 2024 Oct. MedComm (2020). 2024. PMID: 39309696 Free PMC article.
-
Directly targeting PRDM16 in thermogenic adipose tissue to treat obesity and its related metabolic diseases.Front Endocrinol (Lausanne). 2024 Sep 16;15:1458848. doi: 10.3389/fendo.2024.1458848. eCollection 2024. Front Endocrinol (Lausanne). 2024. PMID: 39351529 Free PMC article. Review.
References
-
- Kuhnisch J, Herbst C, Al-Wakeel-Marquard N, Dartsch J, Holtgrewe M, Baban A, Mearini G, Hardt J, Kolokotronis K, Gerull B, Carrier L, Beule D, Schubert S, Messroghli D, Degener F, Berger F, Klaassen S. Targeted panel sequencing in pediatric primary cardiomyopathy supports a critical role of TNNI3. Clin Genet 2019;96:549–559. - PubMed
-
- Arndt AK, Schafer S, Drenckhahn JD, Sabeh MK, Plovie ER, Caliebe A, Klopocki E, Musso G, Werdich AA, Kalwa H, Heinig M, Padera RF, Wassilew K, Bluhm J, Harnack C, Martitz J, Barton PJ, Greutmann M, Berger F, Hubner N, Siebert R, Kramer HH, Cook SA, MacRae CA, Klaassen S. Fine mapping of the 1p36 deletion syndrome identifies mutation of PRDM16 as a cause of cardiomyopathy. Am J Hum Genet 2013;93:67–77. - PMC - PubMed
-
- Delplancq G, Tarris G, Vitobello A, Nambot S, Sorlin A, Philippe C, Carmignac V, Duffourd Y, Denis C, Eicher JC, Chevarin M, Millat G, Khallouk B, Rousseau T, Falcon-Eicher S, Vasiljevic A, Harizay FT, Thauvin-Robinet C, Faivre L, Kuentz P. Cardiomyopathy due to PRDM16 mutation: first description of a fetal presentation, with possible modifier genes. Am J Med Genet C Semin Med Genet 2020;184:129–135. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
