

Bhlhe40 limits early IL-10 production from CD4+ T cells during Plasmodium yoelii 17X infection
- PMID: 37843306
- PMCID: PMC10652903
- DOI: 10.1128/iai.00367-23
Bhlhe40 limits early IL-10 production from CD4+ T cells during Plasmodium yoelii 17X infection
Abstract
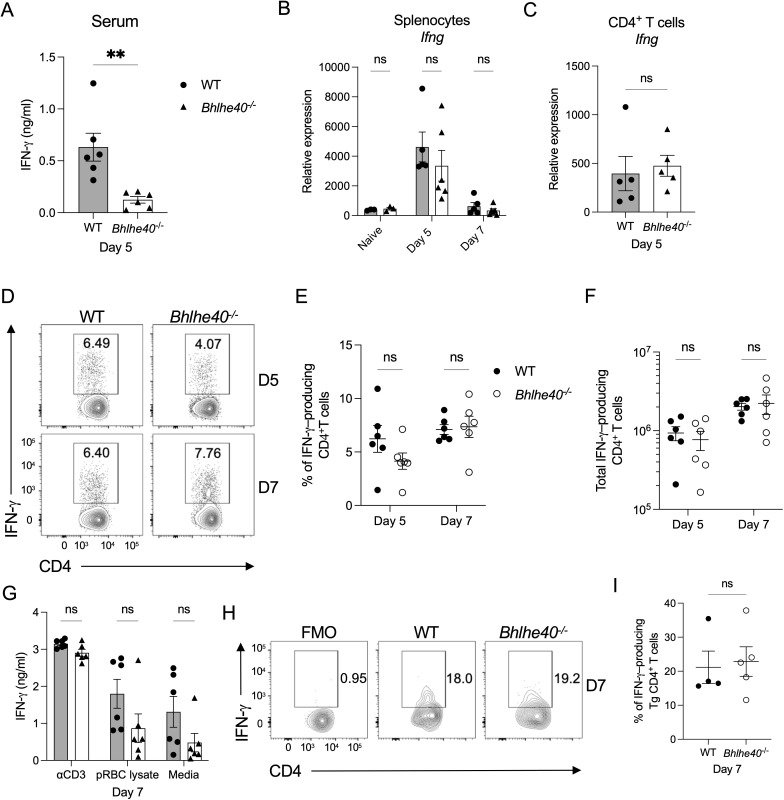
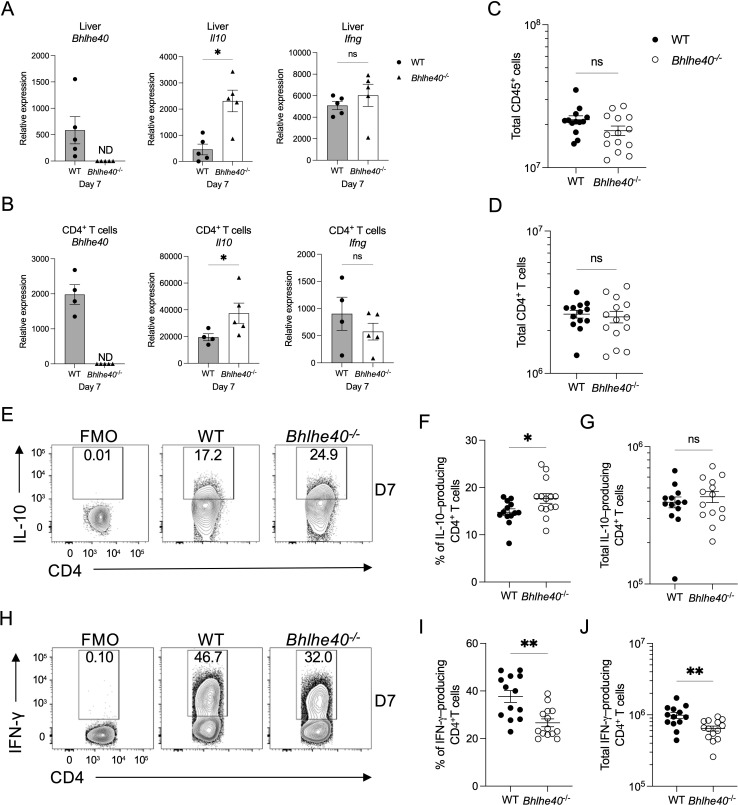
The cytokine IL-10 suppresses T-cell-mediated immunity, which is required to control infection with Plasmodium yoelii. Consequently, IL-10 can delay the time needed to resolve this infection, leading to a higher parasite burden. While the pathways that lead to IL-10 production by CD4+ T cells are well defined, much less is known about the mediators that suppress the expression of this potent anti-inflammatory cytokine. Here, we show that the transcription factor basic helix-loop-helix family member e40 (Bhlhe40) contributes to controlling parasite burden in response to P. yoelii infection in mice. Loss of Bhlhe40 expression in mice results in higher Il10 expression, higher peak parasitemia, and a delay in parasite clearance. The observed phenotype was not due to defects in T-cell activation and proliferation or the humoral response. Nor was it due to changes in regulatory T-cell numbers. However, blocking IL-10 signaling reversed the outcome in Bhlhe40-/ - mice, suggesting that excess IL-10 production limits their ability to control the infection properly. In addition to suppressing Il10 expression in CD4+ T cells, Bhlhe40 can promote Ifng expression. Indeed, IFN-γ production by CD4+ T cells isolated from the liver was significantly affected by the loss of Bhlhe40. Lastly, Bhlhe40 deletion in T cells resulted in a phenotype similar to that observed in the Bhlhe40-/ - mice, indicating that Bhlhe40 expression in T cells contributes to the ability of mice to control infection with P. yoelii.
Keywords: Plasmodium; T cells; cytokines; infectious disease; transcription factors.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Similar articles
-
BHLHE40 drives protective polyfunctional CD4 T cell differentiation in the female reproductive tract against Chlamydia.PLoS Pathog. 2024 Jan 25;20(1):e1011983. doi: 10.1371/journal.ppat.1011983. eCollection 2024 Jan. PLoS Pathog. 2024. PMID: 38271477 Free PMC article.
-
Transcriptional regulator Bhlhe40 works as a cofactor of T-bet in the regulation of IFN-γ production in iNKT cells.Proc Natl Acad Sci U S A. 2016 Jun 14;113(24):E3394-402. doi: 10.1073/pnas.1604178113. Epub 2016 May 25. Proc Natl Acad Sci U S A. 2016. PMID: 27226296 Free PMC article.
-
BHLHE40 Regulates IL-10 and IFN-γ Production in T Cells but Does Not Interfere With Human Type 1 Regulatory T Cell Differentiation.Front Immunol. 2021 Jul 7;12:683680. doi: 10.3389/fimmu.2021.683680. eCollection 2021. Front Immunol. 2021. PMID: 34305917 Free PMC article.
-
Bhlhe40 is an essential repressor of IL-10 during Mycobacterium tuberculosis infection.J Exp Med. 2018 Jul 2;215(7):1823-1838. doi: 10.1084/jem.20171704. Epub 2018 May 17. J Exp Med. 2018. PMID: 29773644 Free PMC article.
-
BHLHE40 Regulates Myeloid Cell Polarization through IL-10-Dependent and -Independent Mechanisms.J Immunol. 2024 Jun 1;212(11):1766-1781. doi: 10.4049/jimmunol.2200819. J Immunol. 2024. PMID: 38683120 Free PMC article.
Cited by
-
BHLHE40 drives protective polyfunctional CD4 T cell differentiation in the female reproductive tract against Chlamydia.PLoS Pathog. 2024 Jan 25;20(1):e1011983. doi: 10.1371/journal.ppat.1011983. eCollection 2024 Jan. PLoS Pathog. 2024. PMID: 38271477 Free PMC article.
-
BHLHE40 drives protective polyfunctional CD4 T cell differentiation in the female reproductive tract against Chlamydia.bioRxiv [Preprint]. 2023 Nov 5:2023.11.02.565369. doi: 10.1101/2023.11.02.565369. bioRxiv. 2023. Update in: PLoS Pathog. 2024 Jan 25;20(1):e1011983. doi: 10.1371/journal.ppat.1011983. PMID: 37961221 Free PMC article. Updated. Preprint.
References
-
- Crompton PD, Moebius J, Portugal S, Waisberg M, Hart G, Garver LS, Miller LH, Barillas-Mury C, Pierce SK. 2014. Malaria immunity in man and mosquito: insights into unsolved mysteries of a deadly infectious disease. Annu Rev Immunol 32:157–187. doi:10.1146/annurev-immunol-032713-120220 - DOI - PMC - PubMed
-
- Lyke KE, Burges R, Cissoko Y, Sangare L, Dao M, Diarra I, Kone A, Harley R, Plowe CV, Doumbo OK, Sztein MB. 2004. Serum levels of the proinflammatory cytokines interleukin-1 beta (IL-1beta), IL-6, IL-8, IL-10, tumor necrosis factor alpha, and IL-12(p70) in Malian children with severe Plasmodium falciparum malaria and matched uncomplicated malaria or healthy controls. Infect Immun 72:5630–5637. doi:10.1128/IAI.72.10.5630-5637.2004 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
