

The Influence of Human IgG Subclass and Allotype on Complement Activation
- PMID: 37843500
- PMCID: PMC10656437
- DOI: 10.4049/jimmunol.2300307
The Influence of Human IgG Subclass and Allotype on Complement Activation
Abstract
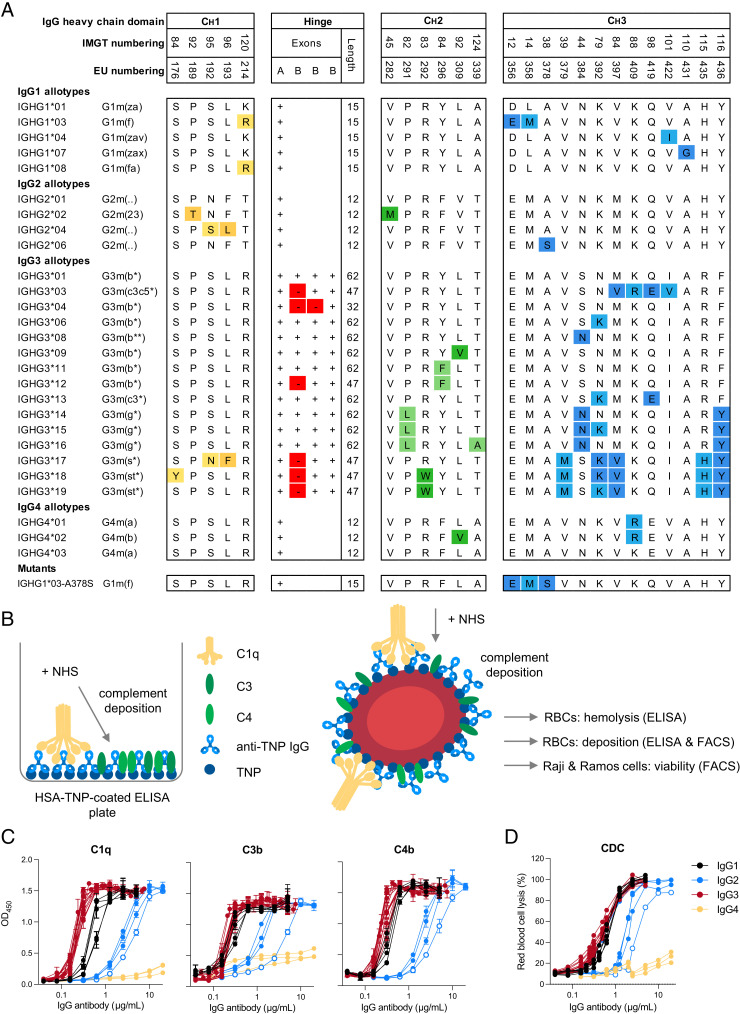
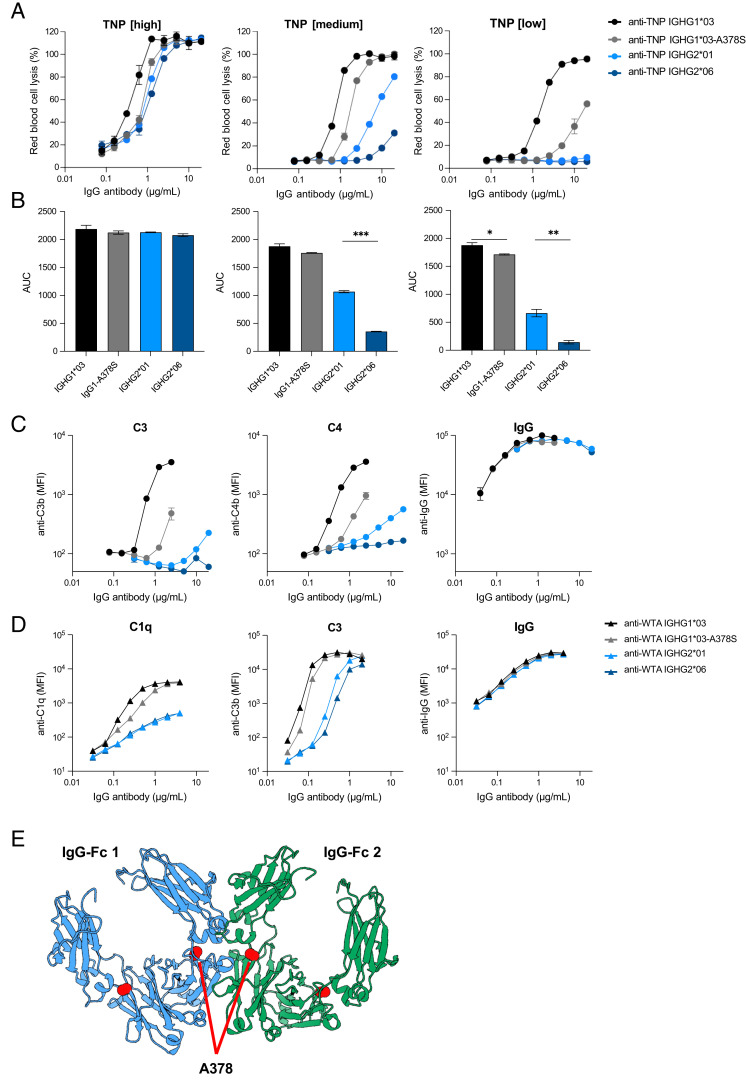
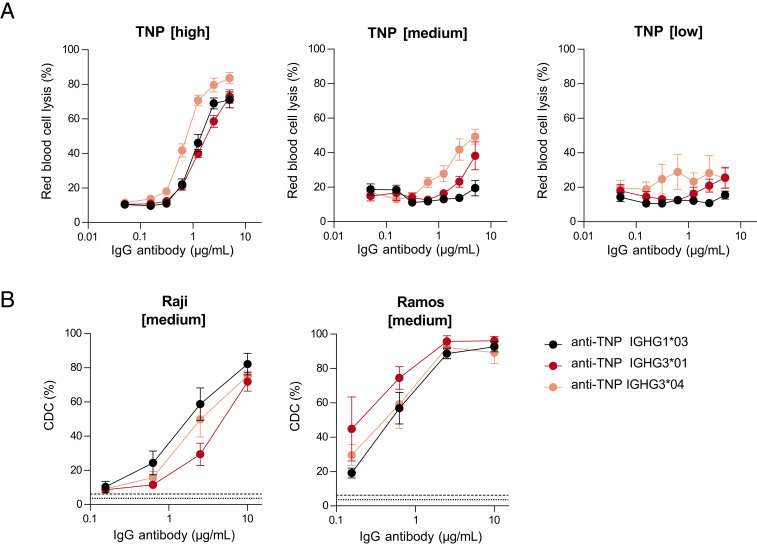
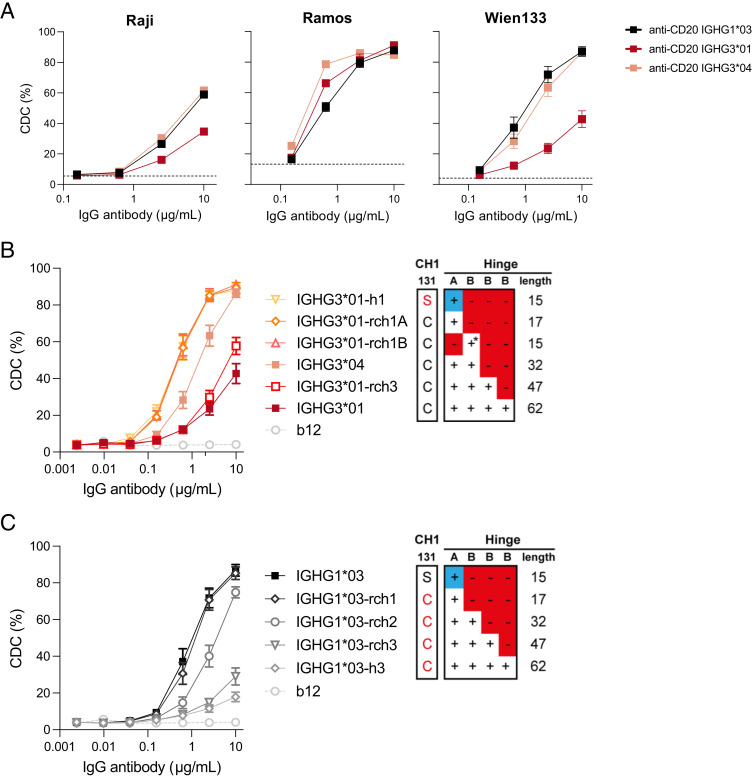
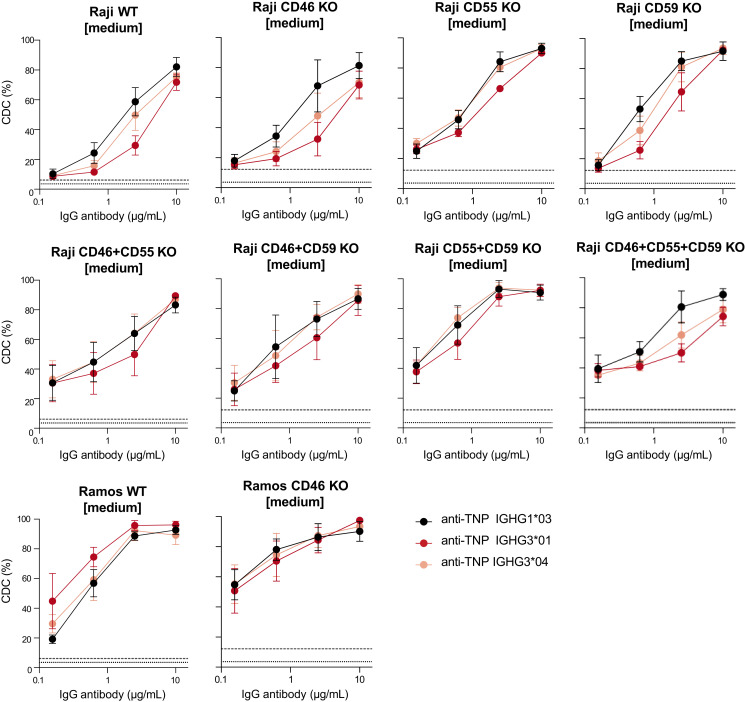
Complement activation via the classical pathway is initiated when oligomeric Igs on target surfaces are recognized by C1 of the complement cascade. The strength of this interaction and activation of the complement system are influenced by structural variation of the Ab, including Ab isotype, subclass, and glycosylation profile. Polymorphic variants of IgG have also been described to influence Fc-dependent effector functions. Therefore, we assessed complement binding, deposition, and complement-dependent cytotoxicity (CDC) of 27 known IgG allotypes with anti-trinitrophenyl specificity. Differences between allotypes within subclasses were minor for IgG1, IgG3, and IgG4 allotypes, and more substantial for IgG2. Allelic variant IGHG2*06, containing a unique serine at position 378 in the CH3 domain, showed less efficient complement activation and CDC compared with other IgG2 polymorphisms. We also observed variable cell lysis between IgG1 and IgG3, with IgG3 being superior in lysis of human RBCs and Ramos cells, and IgG1 being superior in lysis of Raji and Wien133 cells, demonstrating that a long-standing conundrum in the literature depends on cellular context. Furthermore, we compared IgG1 and IgG3 under different circumstances, showing that Ag density and Ab hinge length, but not complement regulators, define the context dependency of Ab-mediated CDC activity. Our results point toward a variation in the capacity of IgG subclasses to activate complement due to single amino acid changes and hinge length differences of allotypes to activate complement, which might give new insights on susceptibility to infectious, alloimmune, or autoimmune diseases and aid the design of Ab-based therapeutics.
Copyright © 2023 by The American Association of Immunologists, Inc.
Conflict of interest statement
A.F.L., M.M.M. and J.S. are former employees of Genmab and own stock and/or warrants. T.D. and S.W.d.T. were awarded Genmab-sponsored research funding. The other authors have no financial conflicts of interest.
Figures
References
-
- Strasser, J., de Jong R. N., Beurskens F. J., Wang G., Heck A. J. R., Schuurman J., Parren P. W. H. I., Hinterdorfer P., Preiner J.. 2019. Unraveling the macromolecular pathways of IgG oligomerization and complement activation on antigenic surfaces. Nano Lett. 19: 4787–4796. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
