

Resolution in cryogenic solid state NMR: Challenges and solutions
- PMID: 37847566
- PMCID: PMC11184935
- DOI: 10.1002/pro.4803
Resolution in cryogenic solid state NMR: Challenges and solutions
Abstract
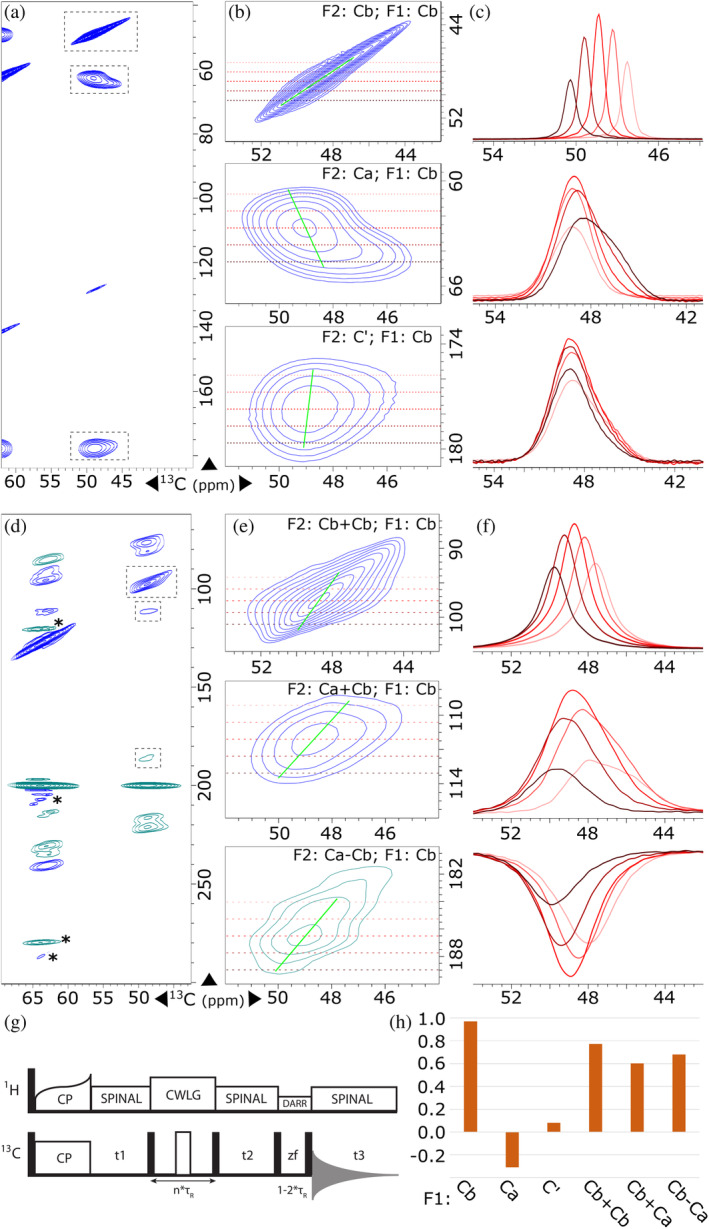
NMR at cryogenic temperatures has the potential to provide rich site-specific details regarding biopolymer structure, function, and mechanistic intermediates. Broad spectral lines compared with room temperature NMR can sometimes present practical challenges. A number of hypotheses regarding the origins of line broadening are explored. One frequently considered explanation is the presence of inhomogeneous conformational distributions. Possibly these arise when the facile characteristic motions that occur near room temperature become dramatically slower or "frozen out" at temperatures below the solvent phase change. Recent studies of low temperature spectra harness the distributions in properties in these low temperature spectra to uncover information regarding the conformational ensembles that drive biological function.
Keywords: DNP; cryogenic NMR; dynamic nuclear polarization; magnetic resonance; protein dynamics; solid state NMR.
© 2023 The Protein Society.
Figures






References
-
- Bajaj VS, van der Wel PCA, Griffin RG. Observation of a low‐temperature, dynamically driven structural transition in a polypeptide by solid‐state NMR spectroscopy observation of a low‐temperature, dynamically driven structural transition in a polypeptide by solid‐state NMR. 2009;125:118–128. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
