

Serotonin reduction in post-acute sequelae of viral infection
- PMID: 37848036
- PMCID: PMC11227373
- DOI: 10.1016/j.cell.2023.09.013
Serotonin reduction in post-acute sequelae of viral infection
Abstract
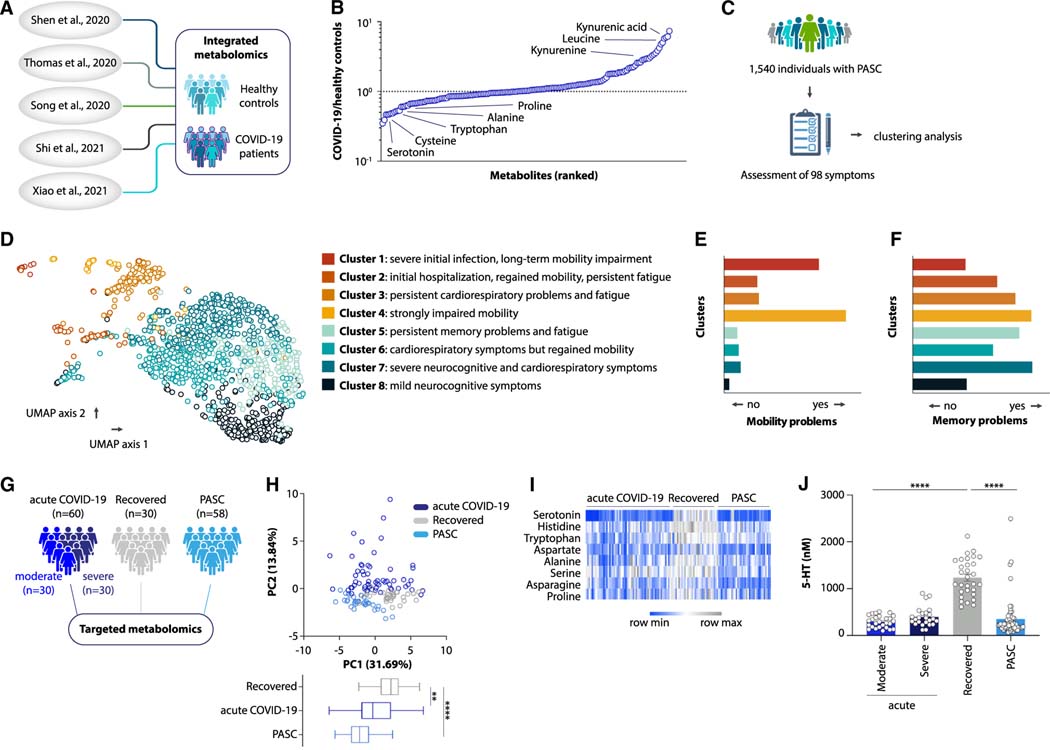
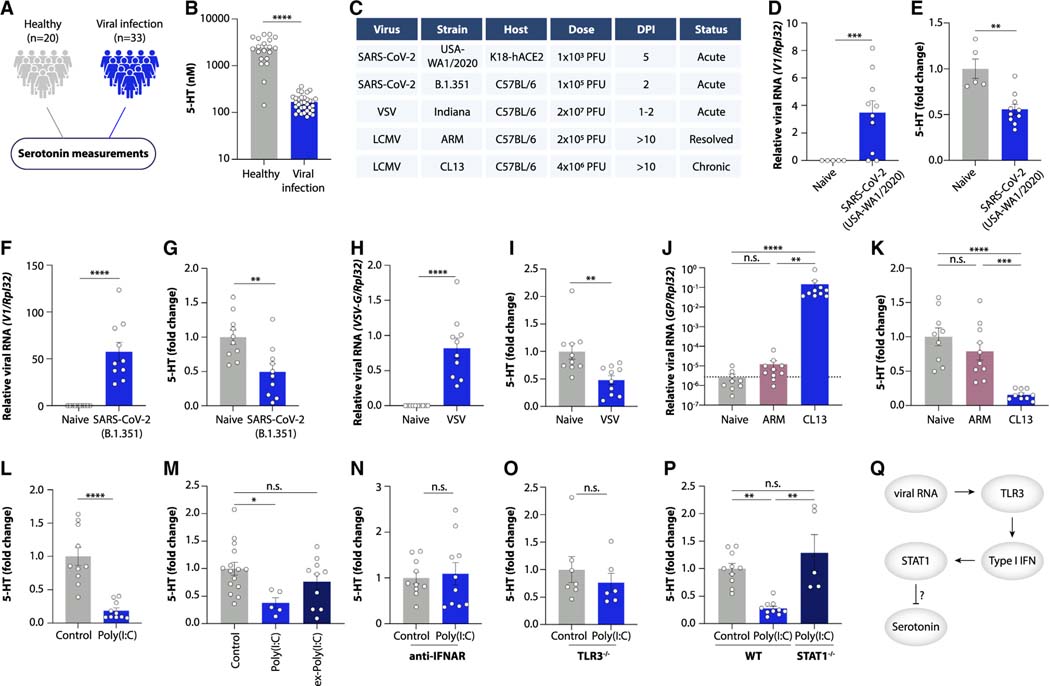
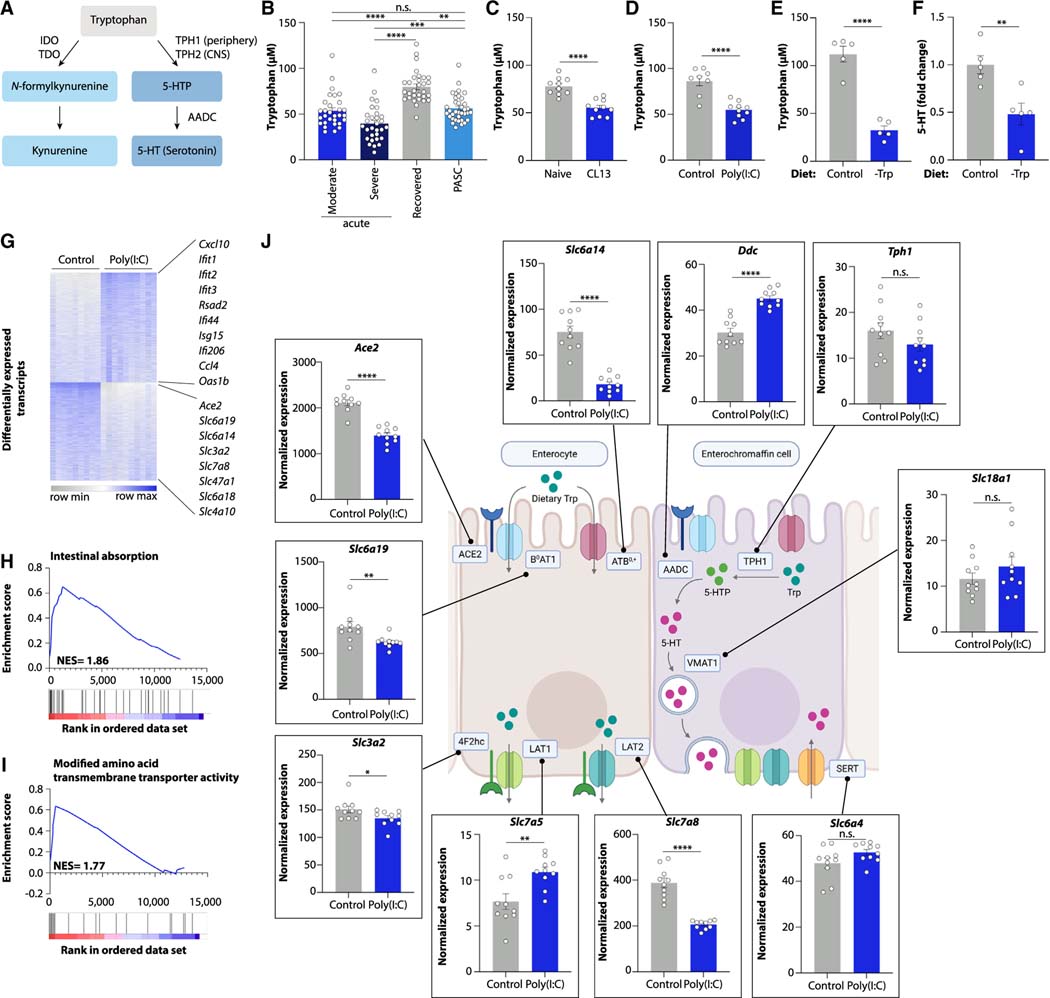
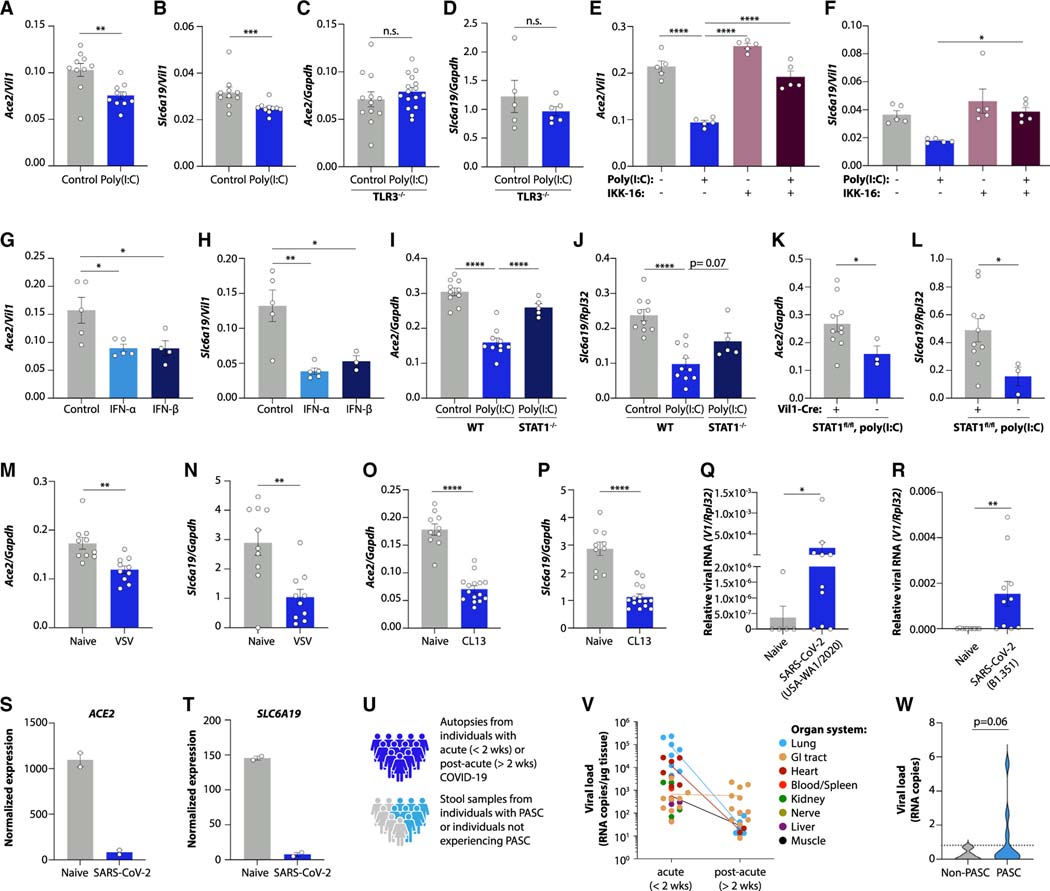
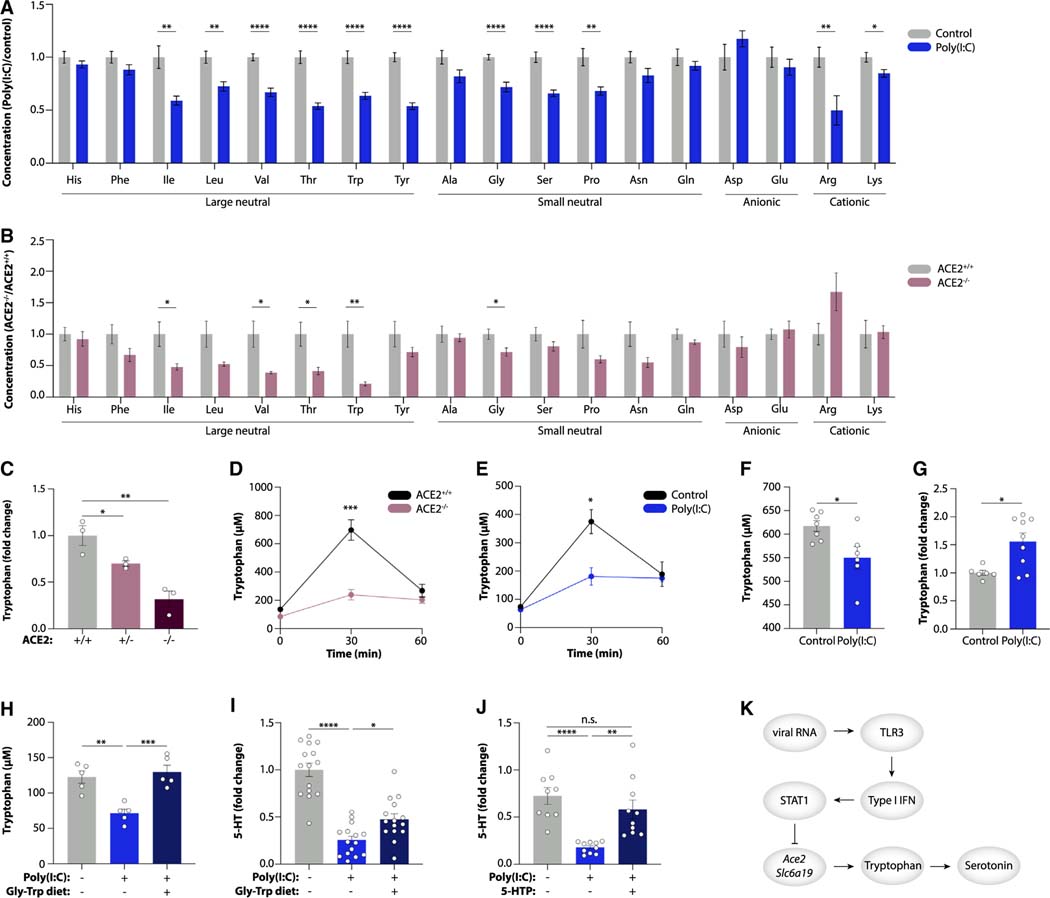
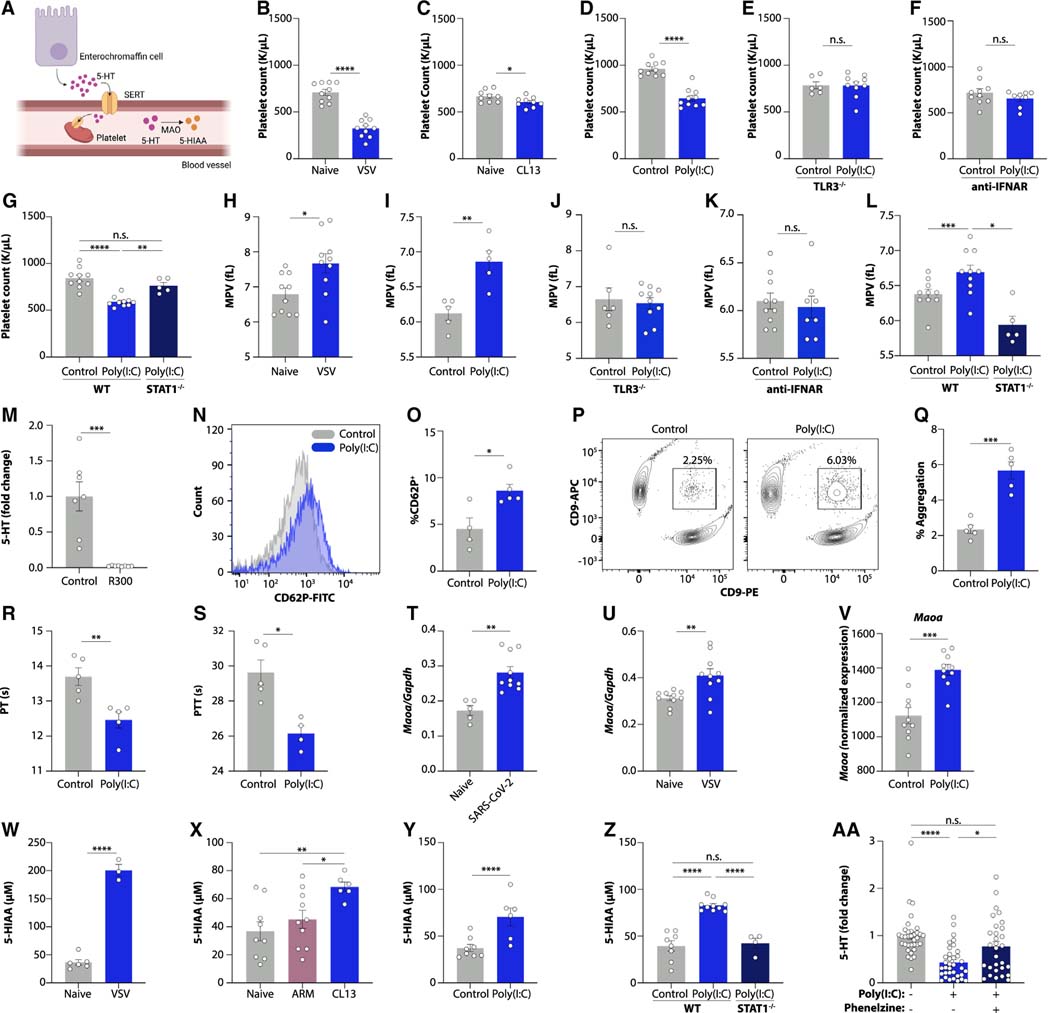
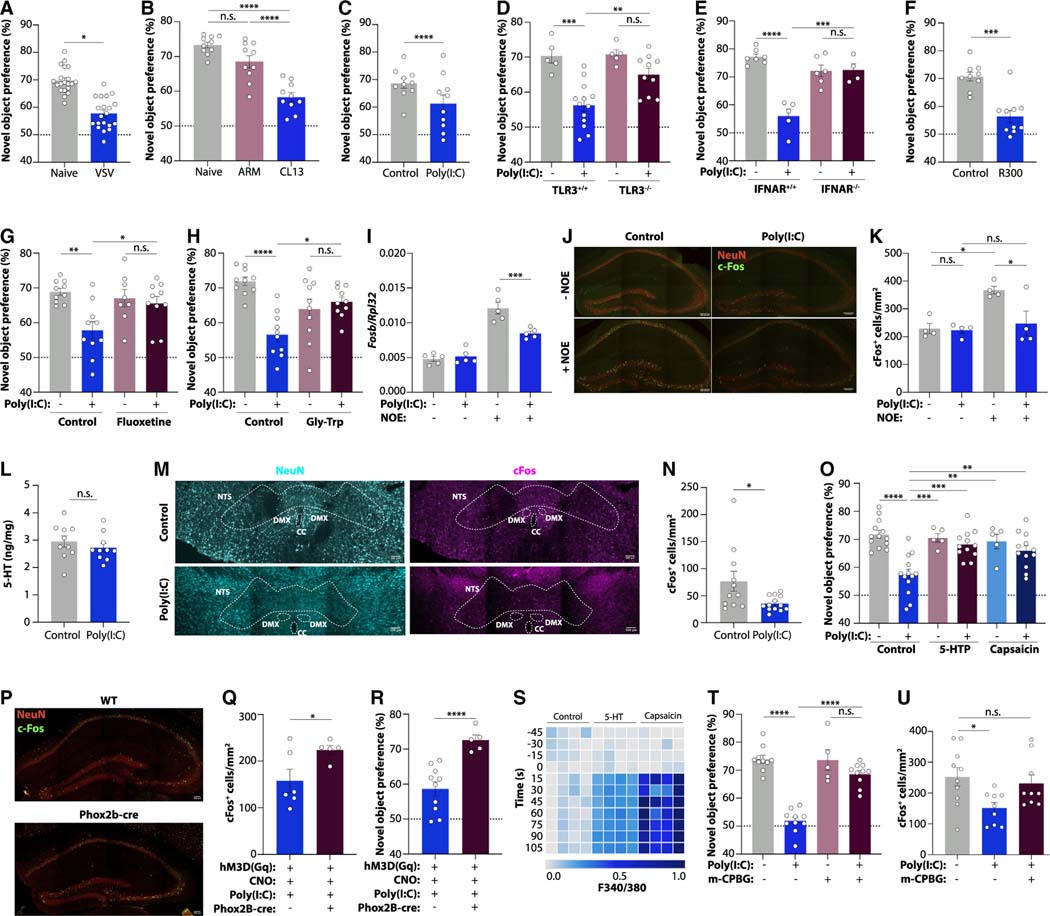
Post-acute sequelae of COVID-19 (PASC, "Long COVID") pose a significant global health challenge. The pathophysiology is unknown, and no effective treatments have been found to date. Several hypotheses have been formulated to explain the etiology of PASC, including viral persistence, chronic inflammation, hypercoagulability, and autonomic dysfunction. Here, we propose a mechanism that links all four hypotheses in a single pathway and provides actionable insights for therapeutic interventions. We find that PASC are associated with serotonin reduction. Viral infection and type I interferon-driven inflammation reduce serotonin through three mechanisms: diminished intestinal absorption of the serotonin precursor tryptophan; platelet hyperactivation and thrombocytopenia, which impacts serotonin storage; and enhanced MAO-mediated serotonin turnover. Peripheral serotonin reduction, in turn, impedes the activity of the vagus nerve and thereby impairs hippocampal responses and memory. These findings provide a possible explanation for neurocognitive symptoms associated with viral persistence in Long COVID, which may extend to other post-viral syndromes.
Keywords: Long COVID; PASC; neurocognitive symptoms; platelets; post-viral syndromes; serotonin; thrombocytopenia; type I interferons; vagus nerve; viral persistence.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests E.J.W. is an advisor for Danger Bio, Janssen, New Limit, Marengo, Pluto Immunotherapeutics Related Sciences, Rubius Therapeutics, Santa Ana Bio, Synthekine, and Surface Oncology. E.J.W. is a founder of and holds stock in Surface Oncology, Danger Bio, and Arsenal Biosciences. N.J.M. reports consulting fees from Endpoint Health Inc and AstraZeneca and receives funding from Quantum Leap Healthcare Collaborative outside of the published work.
Figures
Comment in
-
Low serotonin linked to long COVID.Nat Rev Immunol. 2023 Dec;23(12):784. doi: 10.1038/s41577-023-00966-7. Nat Rev Immunol. 2023. PMID: 37935992 No abstract available.
References
-
- Pretorius E, Vlok M, Venter C, Bezuidenhout JA, Laubscher GJ, Steenkamp J, and Kell DB (2021). Persistent clotting protein pathology in Long COVID/Post-Acute Sequelae of COVID-19 (PASC) is accompanied by increased levels of antiplasmin. Cardiovasc. Diabetol. 20, 172. 10.1186/s12933-021-01359-7. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI158013/AI/NIAID NIH HHS/United States
- P30 AR069589/AR/NIAMS NIH HHS/United States
- R01 HL137915/HL/NHLBI NIH HHS/United States
- P30 AI045008/AI/NIAID NIH HHS/United States
- DP2 AG067511/AG/NIA NIH HHS/United States
- U19 AI082630/AI/NIAID NIH HHS/United States
- P30 ES013508/ES/NIEHS NIH HHS/United States
- P30 CA016520/CA/NCI NIH HHS/United States
- P01 AI108545/AI/NIAID NIH HHS/United States
- R01 AI141003/AI/NIAID NIH HHS/United States
- T32 AI141393/AI/NIAID NIH HHS/United States
- P01 CA210944/CA/NCI NIH HHS/United States
- R01 AA029859/AA/NIAAA NIH HHS/United States
- K23 AI157875/AI/NIAID NIH HHS/United States
- R01 HL137006/HL/NHLBI NIH HHS/United States
- F31 HL160065/HL/NHLBI NIH HHS/United States
- R01 DK123733/DK/NIDDK NIH HHS/United States
- P30 DK050306/DK/NIDDK NIH HHS/United States
- T32 AI060530/AI/NIAID NIH HHS/United States
- R01 AI155577/AI/NIAID NIH HHS/United States
- U19 AI149680/AI/NIAID NIH HHS/United States
- T32 AI055400/AI/NIAID NIH HHS/United States
- L30 AI147159/AI/NIAID NIH HHS/United States
- P30 DK019525/DK/NIDDK NIH HHS/United States
- R35 HL161196/HL/NHLBI NIH HHS/United States
- DP2 AG067492/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
